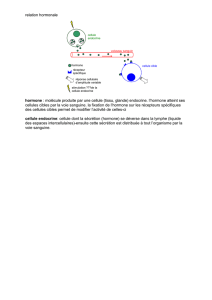
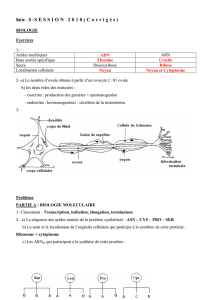
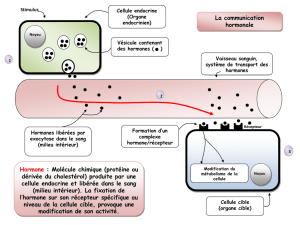
La neurogenèse chez un insecte adulte est sous - iPubli

S MINI-SYNTHÈSE
JO:
L
de

A.

re
1
vo juinuillet
RÉFÉRENCES
Strambi
Colette Sambi
Myriam Cayre
Cnrs chin Josh-Aii
Mal dex 9, F n.
TIRÉS A PART
1
/
3
100%