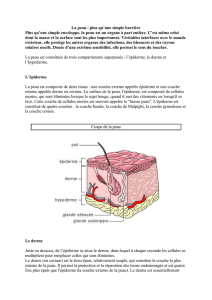
La peau : un des profils autre modèle pour étudier la - iPubli

858
cine/sciences 1992 ; 8 : 858-62
La peau : un
des profils autre modèle pour étudier la formation
morphogénétiques chez les vertébrés
1
4.4,

a
b
a
�
859

860
:
T
b
2.
*GLOSSAIRE *

1
1
1'
?
l.
/
�
�
3.
(a).
1
1
•
Daniee Dhouaiy
N.
861

862
1
/
5
100%