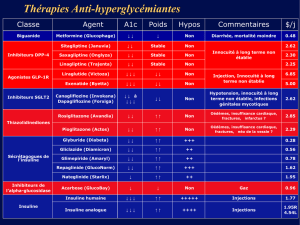
Biologie du diabète.

Médecine Nucléaire - Imagerie fonctionnelle et métabolique - 2001 - vol.25 - n°2 55
G. Baudin
Résumé
Les glucides occupent une place majeure dans le métabolisme énergétique de l’orga-
nisme : ils partagent avec les lipides, et en étroite corrélation avec eux, le rôle de stockage et
d’utilisation de l’énergie.
Deux hormones jouent un rôle central dans la régulation de ces métabolismes : l’insuline
et le glucagon.
Les aspects biochimiques du métabolisme du glucose, de l’insuline et du glucagon sont
rappelés afin d’introduire l’étude analytique et clinique des diabètes sucrés.
Métabolisme glucidique / Insuline / Glucagon
Correspondance : Gérard Baudin - Laboratoire Radio Immunologie - Centre Hospitalier Universitaire - 30029 Nîmes Cedex
Tél : 04 66 68 32 47 - Fax : 04 66 68 32 85
Biologie du diabète.
Gérard Baudin Laboratoire de Radio Immunologie - CHU - Nîmes.
ðLes glucides occupent une place
majeure dans le métabolisme éner-
gétique : des 4 composants organi-
ques classiques (glucides, lipides,
protéines et nucléotides), les gluci-
des partagent avec les lipides, le rôle
LE MÉTABOLISME GLUCIDIQUE :
RAPPELS BIOCHIMIQUES
de stockage de l’énergie, ce qu’ils font
sous forme de polysaccharides.
Les glucides sont apportés à l’orga-
nisme sous une forme le plus géné-
ralement complexe (saccharose/
amidon) et ils sont absorbés au ni-
veau de l’intestin après avoir été scin-
dés en oses simples par des osidases
du suc digestif.
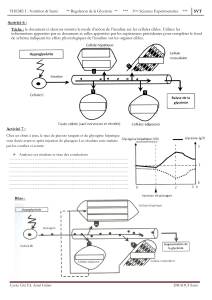
Rôle essentiel du foie
(figure 1figure 1
figure 1figure 1
figure 1)
Le glucose est conduit au foie par la
veine porte et là, est transformé en
glucose-6-phosphate (G6P), qui peut
suivre 5 voies différentes suivant les
besoins de l’organisme, réglés par des
mécanismes hormonaux.

Biologie du diabète
Médecine Nucléaire - Imagerie fonctionnelle et métabolique - 2001 - vol.25 - n°2
56
Intestin
Glucides
Glucose
Veine Porte Glucose circulant
Hépatocyte
Glycogène (Glucose 6 phosphatase)
G 1 P
G 6 P
Neoglucogenèse
Glycolyse Voie des pentoses
(NADPH / ribose)
Ac. Pyruvique
Ac. Aminés
Acétyl CoA
Ac. Nucleiques
(Cycle de Krebs)
Ac. Gras
(Lipides)
CO2 + H2O + énergie Corps cétoniques
Figure 1. Métabolisme hépatique des glucides.
1. Hydrolyse en glucose et phosphate
par la glucose-6-phosphatase (quasi
exclusivité de l’hépatocyte - un peu
dans le rein -) produisant du glucose
circulant distribué aux cellules.
2. Transformation en énergie par l’in-
termédiaire de la glycolyse (figurefigure
figurefigure
figure
22
22
2) : une molécule de glucose donne
2 acides pyruviques eux même trans-
formés en acides oxalo-acétiques
(AOA) et acétyl coenzyme A (acétyl
CoA) : ces deux molécules donnent
naissance au cycle de Krebs qui pro-
duira CO2 + H2O + NAD réduit et
FAD réduit. Ces nucléotides pyridi-
niques réduits donneront, par l’inter-
médiaire de la "chaîne respiratoire",
des molécules d’ATP concourant à la
distribution de l’énergie.
Une molécule de glucose permet
ainsi la fabrication de 22 liaisons éner-
gétiques sous forme de nucléoside-
phosphates.
3. Transformation en nucléotides
pyridiniques réduits (ici NADPH) par
la "voie des pentoses-phosphate" : ce
nucléotide est essentiel dans les tis-
sus qui synthétisent des acides gras
et des stéroïdes (cholestérol) à partir
de l’acétyl CoA.
En même temps, cette voie synthé-
tise des pentoses (le D ribose essen-
tiellement), nécessaires à la synthèse
des acides nucléiques.
Enfin, cette voie dégrade les pento-
ses en hexoses qui peuvent rejoin-
dre la glycolyse.
4. Stockage dans les cellules hépati-
ques et musculaires par polyconden-
sation en glycogène : le "carrefour"
étant ici le glucose-1-phosphate
(G1P). La glycogénolyse servant à ali-
menter
-dans le muscle, la production
d’énergie (glycogèneàG1PàG6Pà
glycolyse),
-dans le foie : la production d’éner-
gie et la régulation de la glycémie (gly-
cogèneàG1PàG6Pàglucose libre).
5. Stockage sous forme de lipides lors-
que toutes les voies précédentes sont
saturées : c’est l’acétyl CoA et l’acide
citrique du cycle de Krebs qui vont
initier cette voie de fabrication des
acides gras.
Ces mêmes acides gras servent de
"réserve" puisque leur dégradation va
fournir de l’acétyl CoA qui pourra
être utilisé dans le cycle de Krebs.
Le fLe f
Le fLe f
Le foie est donc l’oroie est donc l’or
oie est donc l’oroie est donc l’or
oie est donc l’organe prgane pr
gane prgane pr
gane principalincipal
incipalincipal
incipal
régulateur du métabolisme glucidi-régulateur du métabolisme glucidi-
régulateur du métabolisme glucidi-régulateur du métabolisme glucidi-
régulateur du métabolisme glucidi-
queque
queque
que : en effet il est le seul organe qui
laisse "sortir" le glucose.
Les cellules périphériques
(figure 3figure 3
figure 3figure 3
figure 3)
Les autres cellules utilisent le glucose
dans un but de synthèse énergétique
(ATP), de synthèse de réserves (gly-
cogène, lipides) ou de synthèse de
glycoprotéines.
- Lors de l’utilisation musculaire, les
myocytes utilisent l’énergie (ATP –
Créatine-phosphate) et dégradent
leurs réserves (glycogène) : l’acide
lactique, déchet produit par anaéro-
biose, est reconduit au foie où il
pourra être retransformé en G6P
(néoglucogenèse) (figure 4figure 4
figure 4figure 4
figure 4). Cette
Figure 2. Glycolyse.
G 6 P
2 Acides pyruviques
AOA Acétyl CoA
H2O CO2
NADH2
FADH2
(Chaîne respiratoire)
ATP
H2O
Cycle de
Krebs

Médecine Nucléaire - Imagerie fonctionnelle et métabolique - 2001 - vol.25 - n°2 57
G. Baudin
néoglucogénèse prendra le relais, par
l’intermédiaire du foie, lorsque la ré-
serve en glycogène sera insuffisante.
L’alanine et les autres acides aminés
glucoformateurs (et donc les protéi-
nes) ou le glycérol (donc les lipides)
seront fournisseurs de sucres.
Sang
Glucose
Glyco
g
ène (muscle)
Proteo
g
lycanes
G 1 P Glycoproteines
G 6 P
Glycolyse Voie des pentoses
Ac. Pyruvique
Acétyl CoA Ac. Lactique
Ener
g
ie
Cellule périphérique
Retour au foie (neo
g
luco
g
enèse)
Sang
Figure 3. Métabolisme périphérique des glucides.
G 6 P
Acides Aminés glucoformateurs Acide lactique
Sang
Alanine Glucose Acide lactique
Acides Aminés G 6 P Acide lactique
Proteines Energie
Cellule périphérique
Figure 4. Néoglycogenèse.
- Une dernière voie intéressante est
la cétogenèsecétogenèse
cétogenèsecétogenèse
cétogenèse (figure 5figure 5
figure 5figure 5
figure 5) : c’est une
voie mineure de dégradation de
l’acétyl CoA, uniquement hépatique.
Lors du catabolisme des acides gras,
une quantité importante d’acétyl CoA
est formée et doit donc être métabo-
lisée.
Pour cela, les acétyl CoA se "conden-
sent" en corps cétoniques (acide
acétylacétique et acide β OH butyri-
que), quittent l’hépatocyte et sont
retrouvés au niveau des cellules pé-
riphériques où ils sont alors retrans-
formés en acétyl CoA qui sera utilisé
comme source énergétique (cette
utilisation étant stimulée par l’insu-
line).
On comprend donc, à ce niveau, les
phénomènes physio-pathologiques :
-La cétonémie est normalement fai-
ble (noter que les corps cétoniques
inhibent la glycolyse et la lipolyse ;
ce qui conduit à leur utilisation éner-
gétique préférentielle).
-Elle augmente au cours du jeûne
glucidique : utilisation accrue des
acides gras avec insuffisance d’utili-
sation au niveau hépatique (déficit en
AOA) donc passage sanguin, c’est la
cétoacidose du jeûne.
-Au cours d’un diabète de type 1, il
y aura :
. Déficit en insuline : diminution de
l’entrée du glucose dans la cellule.
. Arrêt de la glycolyse normale donc
néoglucogenèse accrue, d’où utilisa-
tion de l’AOA et déficit en ce méta-
bolite.
. Catabolisme des acides gras accru
dans les hépatocytes, d’où accumu-
lation d’acétyl CoA.
. En l’absence d’AOA, le métabo-
lisme de l’acétyl CoA se fera donc par
la voie des corps cétoniques.
. Pas d’utilisation périphérique des
corps cétoniques par déficit en insu-
line, d’où cétoacidose très importante
(coma acidocétosique).
Catabolisme des Acides Gras
Acétyl CoA
Corps cétoniques Cycle de Krebs
Tissus périphériques
Acétyl CoA Utilisation
Figure 5. Cétogenèse.

Biologie du diabète
Médecine Nucléaire - Imagerie fonctionnelle et métabolique - 2001 - vol.25 - n°2
58
INSULINE ET GLUCAGON :
RAPPELS BIOCHIMIQUES
Plusieurs hormones participent au
contrôle de l’équilibre glycémique.
Le rôle majeur est toutefois joué par
les hormones pancréatiques que sont
l’insuline et le glucagon.
Ces hormones sont sécrétées dans la
portion endocrine du pancréas, les
îlots de Langerhans, qui ne consti-
tuent que 1%, en poids, de cet organe.
Disséminés dans le tissu exocrine, les
îlots assurent la sécrétion de l’insu-
line par leurs cellules β et du glucagon
par leurs cellules α.
L’insuline
Structure
L’insuline, quelle que soit l’espèce,
est constituée de 2 chaînes.
Les insulines des mammifères ont
une structure primaire très proche,
l’insuline de porc ne différant de l’in-
suline humaine que par 1 acide aminé
et l’insuline de bœuf par 2 acides
aminés seulement.
La molécule d’insuline est constituée
de deux chaînes polypeptidiques :
une de 21 acides aminés (chaîne A)
et une de 30 acides aminés (chaîneB).
Ces deux chaînes sont reliées par
deux ponts disulfure A7-B7 et A20-B19
(figure 6figure 6
figure 6figure 6
figure 6).
La configuration tridimensionnelle
des insulines des différentes espèces
est très proche, expliquant ainsi que
l’activité biologique des insulines soit
semblable. Le monomère, forme
circulante principale, a un poids mo-
léculaire d’environ 6000.
Biosynthèse
L’insuline est formée à partir d’un pré-
curseur, la proinsuline de poids mo-
léculaire d’environ 9000.
La proinsuline est monocaténaire : les
deux chaînes de l’insuline étant re-
liées, de l’extrémité C terminale de
la chaîne B à l’extrémité N terminale
de la chaîne A, par un peptide, le pep-
tide de connexion ou C peptide (fi-fi-
fi-fi-
fi-
gure 7gure 7
gure 7gure 7
gure 7).
Figure 6. Structure de l'insuline.
Figure 7. Structure de la proinsuline.

Médecine Nucléaire - Imagerie fonctionnelle et métabolique - 2001 - vol.25 - n°2 59
G. Baudin
La transformation de proinsuline en
insuline dans l’appareil de Golgi et
dans les granules de transport jusqu’à
la membrane cellulaire (demi-pé-
riode d’environ 60 minutes) fait in-
tervenir des enzymes à activité tryp-
sine et carboxypeptidase et conduit
à la formation de quantités équimo-
laires d’insuline et de C peptide.
La glycémie est le stimulus principal
de la biosynthèse et de la sécrétion
d’insuline.
Secrétion
L’insuline et le C peptide des granu-
les sont libérés par exocytose : ce
mode de sécrétion - le principal - est
fonction des stimuli primaires (gly-
cémie surtout) et secondaires (hor-
mones).
Une sécrétion de base existe, en de-
hors de tout stimulus exogène.
En réponse aux stimuli, la sécrétion
est biphasique : d’abord une élévation
en moins d’une minute représentant
l’insuline stockée dans le comparti-
ment immédiatement mobilisable,
puis, après une diminution, une
deuxième sécrétion plus lente repré-
sentant de l’insuline stockée mais
aussi une insuline nouvellement syn-
thétisée.
Les stimuli influençant la sécrétion
d’insuline sont de diverses origines.
-Des métabolites : le glucose, la plu-
part des acides aminés (arginine sur-
tout), les acides gras et les corps cé-
toniques dans une moindre mesure
stimulent la secrétion.
-Des hormones : hormones gastro-
intestinales (gastrine, secrétine, gluca-
gon), hGH, glucocorticoïdes, oestro-
gènes, progestérone stimulent la
secrétion.
-Des neuro-médiateurs : les agents
cholinergiques stimulent la sécrétion
et les catécholamines l’inhibent.
Les α bloquants stimulent donc la
sécrétion et les β bloquants la dépri-
ment.
Circulation et catabolisme
Insuline, proinsuline et C peptide cir-
culent sous forme libre, l’insuline
sous forme de monomère.
La proportion de proinsuline circu-
lante peut représenter jusqu’à 20 %
de l’immunoréactivité insulinique
totale et peut-être considérablement
accrue dans certains états pathologi-
ques.
La demi-vie de l’insuline dans le sec-
teur vasculaire sanguin est brève -
environ 5 minutes -:
-le foie, organe important d’action
et de dégradation, prélève 50 % de
l’insulinémie en un passage - celle-ci
est donc nettement plus élevée dans
la veine porte -.
-le rein prélève, lui, 40 % de l’insuli-
némie - l’insuffisance rénale se tra-
duit donc par une demi-vie plasmati-
que de l’insuline augmentée -.
Presque tous les tissus de l’orga-
nisme utilisent et dégradent l’insuline,
le foie et le rein étant les deux sites
majeurs.
La demi-vie de la proinsuline est d’en-
viron 20 minutes : sa dégradation ne
s’accompagne pas de conversion en
insuline ; c’est d’autre part un inhibi-
teur compétitif de la dégradation de
l’insuline.
Le C peptide n’est pas dégradé et est
éliminé sans modification dans
l’urine.
Mode d’action
L’action cellulaire de l’insuline
est médiée par un récepteur - le ré-
cepteur de l’insuline - à activité tyro-
sine-kinase. Ce récepteur, de type 2,
est un tétramère α2 β2.
La liaison extra-cellulaire de l’hor-
mone au récepteur conduit à l’auto-
phosphorylation du domaine intra-
cellulaire. Les récepteurs ainsi activés
sont alors reconnus, grâce à leurs
domaines SH2, par les protéines ci-
bles. Ces protéines, phosphorylées,
jouent alors leur rôle dans la signali-
sation cellulaire.
Actions physiologiques
L’insuline exerce des effets métabo-
liques multiples : son rôle, par son
effet sur l’homéostasie glycémique,
est une stimulation de la mise en ré-
serve de l’énergie ; le glucagon exerce
un effet inverse de mobilisation des
substrats énergétiques.
-Sur les glucides : l’insuline stimule
la glycogènogenèse (foie, muscle
strié) et inhibe la glycogènolyse et la
néoglucogenèse hépatiques.
-Sur les lipides : l’insuline stimule
la lipogenèse (tissu adipeux, foie) et
inhibe la lipolyse et la cétogenèse
(foie). L’utilisation périphérique des
corps cétoniques est favorisée par
l’insuline : le déficit en insuline favo-
risera donc l’apparition d’une acido-
cétose.
-Sur les protéines : l’insuline stimule
le transport intracellulaire des acides
aminés (tissu adipeux, muscle strié)
et accroît leur incorporation dans les
protéines. La synthèse des protéines
est accrue au niveau des ribosomes
dans le foie et le muscle strié.
L’insuline diminue le catabolisme
protidique.
Le glucagon
Structure
Polypeptide de 29 acides aminés, le
glucagon a très peu varié au cours de
l’évolution : la composition en acides
aminés est similaire chez de nom-
breux mammifères et oiseaux. Son
poids moléculaire est d’environ 3500
(figure 8figure 8
figure 8figure 8
figure 8).
NH2-NH2-
NH2-NH2-
NH2-HIS-SER-GLU-THR-PHE-THR-SER-ASP-TYR-SER-LYS-TYR-LEU-ASP-SER-
1 5 10 15
-ARG-ALA-GLU-ASP-GLU-ASP-PHE-VAL-GLU-TRP-LEU-MET-ASN-THR-COOHCOOH
COOHCOOH
COOH
20 25 29
Figure 8. Structure du glucagon.
 6
6
1
/
6
100%