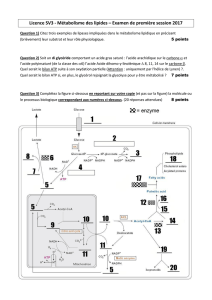
Schéma général des voies lipidiques Les substrats

Master 1 2005-2006
REGULATIONS METABOLIQUES

Schéma global des voies énergétiques à l’exercice

Schéma général des voies glucidiques
La dégradation et la resynthèse du glucose sont les principales
voies énergétiques permettant la réalisation et la continuation de
l’exercice musculaire.
Néanmoins, la resynthèse du glucose peut être faite à partir de
multiples processus biochimiques impliquant divers substrats
et/ou métabolites.
Le schéma général des voies glucidiques montre bien que
l’organisme humain à développé un système biochimique destiné à
synthétiser son substrat énergétique de prédilection : le glucose.
Notons tout de même que plusieurs incertitudes demeurent (ou ont
émergé) quant à cette destinée absolutiste.

MUSCLE CIRCULATION FOIE
Glycogénolyse
Glycogène
G1P
Glycérol 3-P
3-P glycéraldéhyde
Lactate Ac. lactique
Néoglucogenèse
Glucose
Cycle de
Cori
Glucose
Glucose
Acides
aminés Ac. aminés
Ac.Pyruvique
Glycogénolyse
Glycogène
G1P
G6P
Glycolyse
Acétyl-CoA
Ac. lactique
F6P
F1,6B
Ac.Pyruvique
b-Oxydation
KREBS
Oxydation
AGL
TG
AGL
Glycérol
Glycérol
Lipolyse

MUSCLE CIRCULATION FOIE
Glycogène
G1P
Glycérol 3-P
3-P glycéraldéhyde
Lactate Ac. lactique
Glucose
Glucose
Glucose
Acides
aminés Ac. aminés
Ac.Pyruvique
Glycogène
G1P
G6P
Acétyl-CoA
Ac. lactique
F6P
F1,6B
Ac.Pyruvique
KREBS
AGL
TG
AGL
Glycérol
Glycérol
b-oxydation
?
?
?
?
?
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
1
/
42
100%