université paris 6

UNIVERSITÉ PARIS 6
FACULTÉ DE MÉDECINE PIERRE ET
MARIE CURIE
UE2
POLYCOPIÉ DE BIOLOGIE DE LA
REPRODUCTION DE L’ÊTRE HUMAIN
PAES 2012-2013
Polycopié rédigé par Jacqueline Mandelbaum, Catherine Poirot, Célia Ravel et Jean-Pierre
Siffroi

1
OVOGENESE – FOLLICULOGENESE
PR C. POIROT
L'ovaire a une double fonction : une fonction exogène gamétogène, car il assure la
croissance, la maturation puis l'émission de l'ovocyte (gamète femelle) et une fonction
endocrine, puisqu'il synthétise les hormones stéroïdes indispensables à la fonction de
reproduction. C'est entre la puberté et la ménopause que l'ovaire assure cette double fonction.
Le processus qui conduira un follicule donné du stade primordial au stade préovulatoire ou à
l'involution s'appelle la folliculogenèse. Le follicule ovarien est constitué d'un ovocyte
entouré de cellules de la granulosa puis des cellules de la thèque. L'évolution et la maturation
de l'ovocyte sont très liées à la présence des cellules périovocytaires et inversement.
I - OVOGENESE (Figure 1)
L'ovogenèse comporte 3 phases : multiplication, croissance et maturation. Elle commence
pendant la vie fœtale.
a- Phase de multiplication
Les cellules germinatives primordiales de la deuxième poussée embryonnaire donnent
naissance aux ovogonies. Ces ovogonies se divisent, par mitoses simples, du 4ème au 7ème mois
de la vie intra-utérine. Les ovogonies ont, comme les autres cellules de l'organisme, 2N
chromosomes. Cette phase de multiplication est limitée dans le temps. Elle s'achève vers le
7ème mois de la vie intra-utérine. Ces ovogonies, après synthèse d'ADN, vont se transformer
en ovocyte de premier ordre (2N chromosomes, et 2 q (quantité) d'ADN par chromosome [4q
au total pour les 2 chromosomes d'une même paire]).
Cette transformation s'accompagne de 2 phénomènes morphologiques :
- le début de la méiose de la division réductionnelle avec blocage en fin de prophase
de première division : stade dyctié.
- l'organisation autour de chaque ovocyte d'une couche de cellules folliculaires,
constituant ainsi les follicules primordiaux.
b- Phase de croissance
Débute après la naissance, elle est très longue et ne s'achève qu'au moment de la maturation
finale du follicule.
Pendant cette phase de croissance, le volume de l'ovocyte augmente, des synthèses
nucléolaires, nucléaires et cytoplasmiques concourent à donner les éléments nécessaires à la
maturation des ovocytes.
c- Phase de maturation
Elle est définie comme les étapes terminales nécessaires à rendre l'ovocyte fécondable. La
maturation finale de l'ovocyte comprend, une maturation nucléaire consistant en une reprise
de la méiose, une maturation cytoplasmique qui s'accompagne principalement de la synthèse
de protéines spécifiques et de modifications des organites et une maturation membranaire.
Nous ne parlerons ici que de la maturation nucléaire ou méiose.
Le noyau de l'ovocyte I appelé aussi vésicule germinative s'est déplacé à la périphérie du
cytoplasme. Il restera bloqué à ce stade jusqu'à la sécrétion cyclique, entre la puberté et la
ménopause, d'une gonadotrophine hypophysaire induisant l'ovulation, la LH (Luteinizing
Hormone). Trente quatre heures environ après le début de la décharge ovulante de LH, la
première division méiotique, division réductionnelle, s'achève : la prophase I reprend et se
termine, suivie de la métaphase I, de l'anaphase I et de la télophase I. Sur le plan

2
morphologique, ceci se traduit par la formation de 2 cellules de taille très inégale : l'ovocyte
et le premier globulaire polaire. Sur le plan chromosomique, chacune de ces cellules est
constituée de 23 chromosomes avec 2 q d'ADN par chromosome.
L'ovocyte débute la deuxième division de méiose, sans passer par le stade de prophase, et se
bloque à nouveau en métaphase de 2ème division. On parle alors d'ovocyte II. C'est à ce stade
qu'il est expulsé hors de l'ovaire (ovulation) et qu'il est fécondable et éventuellement fécondé.
S'il y a fécondation, l'ovocyte achève sa méiose. L'ovocyte est la seule cellule de l'organisme
qui a besoin d'une autre cellule pour terminer sa différentiation. S'il n'y a pas de fécondation,
il s'atrésie.
En cas de fécondation, l'ovocyte II, bloqué en métaphase de deuxième division de méiose,
reprend sa méiose. L'anaphase de deuxième division a lieu, suivie de la télophase se
manifestant sur le plan morphologique par l'expulsion du deuxième globule polaire. Sur le
plan chromosomique, le noyau ovocytaire a 23 chromosomes avec 1 quantité d'ADN par
chromosome.
II - FOLLICULOGENESE
Elle se situe au niveau du cortex des ovaires. Tous les follicules contiennent un ovocyte
bloqué en fin de prophase de première division. Comme il a été vu au chapitre précédent, c'est
seulement au moment de la décharge ovulante de LH que l'ovocyte reprend sa méiose pour se
bloquer à nouveau en métaphase de 2ème division. L'ovocyte ovulé est bloqué en métaphase de
2ème division de méiose, a donc expulsé son premier globule polaire.
La folliculogenèse débute dès la vie fœtale et s'achève à la ménopause. Durant cet intervalle
de temps, seulement 300 à 400 follicules ovuleront sur le million de petits follicules sains
présents à la naissance.
Les follicules ovariens se décrivent sous différentes formes histologiques qui représentent une
série de stades successifs qui correspondent au cycle folliculaire. (Figure 2)
a- Les follicules primordiaux
C'est le plus petit follicule. Ils sont disposés en périphérie de l'ovaire, sous l'albuginée et sont
répartis en plusieurs couches. Le nombre total de follicules primordiaux est fixé pendant la
vie fœtale. Vers la fin du 7ème mois fœtal, on estime qu'il existe environ 7 millions d'ovocytes
I. Une atrésie survient entre cette date et la naissance si bien qu'on ne dénombre plus qu'un
million d'ovocytes I à cette période. Cette atrésie se poursuit pendant la vie post natale si bien
qu'il n'existe plus qu'environ 300 000 follicules à la puberté et un millier au moment de la
ménopause.
Le diamètre des follicules primordiaux est compris entre 25 et 50 µm de diamètre. Ils sont
constitués par un ovocyte I entouré d'une seule assise de cellules folliculaires, pavimenteuses
qui se présentent, sur une coupe transversale de follicule, sous la forme de 3 à 4 cellules
séparées du stroma par une membrane basale mince : la membrane de Slavjanski.
b- Les follicules primaires
Leur diamètre est compris entre 60 à 80 µm. L'ovocyte I a augmenté de diamètre et est
entouré par une membrane hyaline fine, c'est le début de la mise en place de la zone pellucide,
une assise de cellules cubiques et la membrane de Slavjanski.
c- Les follicules secondaires
Leur taille est de l'ordre de 80 à 180 µm de diamètre. La zone pellucide est maintenant bien
visible. Les cellules folliculaires, disposées en 3 à 4 couches et associées entre elles par des
jonctions de type Gap, constituent la granulosa. A partir d'un certain nombre de couches de
cellules de la granulosa, le stroma, qui entoure le follicule, se modifie pour former la thèque

3
interne. Celle-ci est très vascularisée et renferme des cellules qui présentent les
caractéristiques cytologiques des cellules sécrétrices de stéroïdes. Les follicules secondaires
les plus volumineux qui ont des cellules épithélioïdes sécrétrices dans la thèque interne sont
appelés follicules préantraux.
d- Les follicules antraux ou tertiaires
Ces follicules subissent un accroissement important durant lequel leur taille passe de 180 µm
à plusieurs millimètres. Lors de cette croissance, la morphologie des follicules se modifie
avec apparition de cavités au sein de la couche des cellules folliculaires, ces cavités finissent
par confluer pour former une cavité unique ou antrum. L'ovocyte est refoulé à un des pôles du
follicule. La thèque externe se développe autour de la thèque interne. Cette thèque externe est
riche en tissu conjonctif et en vaisseaux sanguins. Les cellules de la granulosa, plus
nombreuses, groupées autour de l'ovocyte et disposées dans une zone délimitée de la paroi
folliculaire, constituent le cumulus oophorus, qui fait saillie dans l'antrum. Maintenant la
limite entre la thèque interne et la granulosa est définie par une épaisse membrane basale.
e- Le follicule pré-ovulatoire ou mûr ou follicule de De Graaf mesure environ
20 mm de diamètre. Il apparaît comme un "kyste" faisant saillie à la surface de l'ovaire.
L'accumulation de liquide folliculaire entraîne une augmentation de volume de la cavité
antrale.
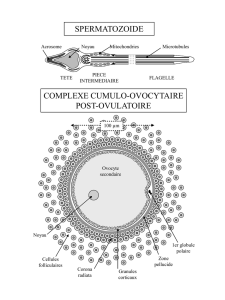
Autour de l'ovocyte, qui a atteint 120 à 150 µm de diamètre, les cellules du cumulus oophorus
les plus proches de l'ovocyte, s'orientent de façon radiaire pour constituer la corona radiata.
Le reste de la granulosa amincie ne comporte que quelques couches de cellules. La thèque
interne est très réduite dans la région qui fait saillie à la surface de l'ovaire.
Lors de la décharge ovulante de LH, dans les heures qui précèdent l'ovulation, les cellules du
cumulus oophorus se transforment profondément. En effet, elles sont dissociées par une
substance mucoïde de telle sorte qu'elles constituent une masse ayant l'aspect d'un petit nuage
de 3 à 4 mm de diamètre.
f- Le follicule au moment de l'ovulation
Sous l'influence de la décharge de LH qui survient à mi-cycle, le follicule présente une série
de modifications :
- Maturation finale de l'ovocyte qui reprend sa méiose, rompt sa vésicule
germinative, expulse son premier globule polaire et se bloque, à nouveau, en
métaphase de 2ème division de méiose. Parallèlement s'effectue la maturation du
cytoplasme.
- Les cellules du cumulus, qui ont la capacité de sécréter des mucopolyssaccharides
se dissocient en entraînant la libération dans la cavité antrale de l'ovocyte II.
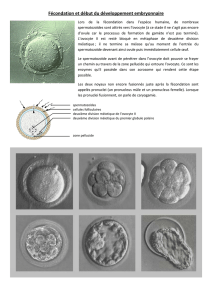
L'ovulation consiste en la rupture du follicule avec libération de l'ovocyte. La rupture de la
paroi folliculaire s'effectue au niveau d'une région appelée le stigma. L'ovocyte II entouré par
la corona radiata et les cellules du cumulus oophorus, est expulsé et capté par le pavillon de la
trompe utérine. En cas de fécondation, l'ovocyte achève sa méiose et expulse son 2ème globule
polaire. Le zygote se divise en gagnant l'utérus. Si l'ovocyte n'est pas fécondé, il dégénère.
g- Le corps jaune
Il correspond à la transformation du follicule après l'ovulation. Ses contours sont irréguliers et
festonnés. La granulosa, sous l'action de la LH, se trouve envahie par des capillaires sanguins
provenant de la thèque interne. Les cellules de la granulosa sont alors modifiées et forment
des cellules du corps jaune sécrétrices de stéroïdes (œstrogènes, et surtout progestérone) ou
cellules lutéales. Les cellules de la thèque interne deviennent les cellules paralutéales.

4
Si l'ovocyte n'est pas fécondé, le corps jaune subit une atrophie en fin de cycle et il ne persiste
qu'une cicatrice fibreuse appelée corpus albicans. S'il y a grossesse, le corps jaune se
maintient et continue son développement. Il devient alors le corps jaune gestatif.
h- L'atrésie folliculaire
La plupart des follicules subissent un processus involutif dénommé atrésie folliculaire. Ce
processus s'observe à tous les stades du développement folliculaire et conduit à l'élimination
de plus de 95% des follicules entrés en phase de croissance. La lésion initiale intéresse
l'ovocyte. Le cytoplasme devient granuleux, renferme de nombreuses enclaves lipidiques, le
noyau se pycnose. Les cellules folliculaires s'altèrent secondairement.
III - HISTOPHYSIOLOGIE ET REGULATION HORMONALE DE LA FOLLICULOGENESE
A- Différentes phases de la folliculogenèse
La folliculogenèse est un phénomène continu dont seuls les 15 derniers jours de
développement est cyclique, cyclicité liée à la régulation hormonale.
Elle peut être divisée en 5 phases :
- la quiescence
- l'entrée en croissance
- le recrutement
- la sélection
- la dominance
Seules les trois dernières étapes sont sous la dépendance des sécrétions des gonadotrophines
hypophysaires donc caractéristiques de la vie génitale active des femmes entre la puberté et la
ménopause. Pendant cette période les ovaires sont à l'origine de sécrétions hormonales
stéroidiennes, sécrétées aussi de façon cyclique (androgènes, œstrogènes et progestérone) et
d'une hormone polypeptidique, l'inhibine.
a- la phase de quiescence
Les follicules quiescents correspondent au pool des follicules primordiaux. Avec le
vieillissement, la population des très petits follicules décroît. Progressivement pour atteindre à
la ménopause un nombre compris entre quelques dizaines et 1000 selon les auteurs.
b- La croissance folliculaire de base
Elle est représentée, au niveau du follicule, par le passage du stade primordial au stade
primaire jusqu'au stade préantral voire antral précoce. Ce passage peut être soit très rapide soit
très long. Il est indépendant de la présence des gonadotrophines d'où son existence avant la
puberté. Cette entrée en croissance est liée à des régulations paracrine/autocrine agissant d'une
façon non encore élucidée. La durée de cette étape n'est pas connue chez la femme.
c- Recrutement, sélection et dominance
Les étapes suivantes correspondent aux 15 derniers jours de la folliculogenèse et sont sous la
dépendance des gonadotrophines. En conséquence, pour expliquer et comprendre les phases
suivantes de la folliculogenèse, il faut aborder sa régulation hormonale.
B- Régulation hormonale des derniers stades de la folliculogenèse
L'ovaire présente une activité cyclique. La durée du cycle ovarien est en moyenne de 28 jours.
Durant ce cycle, on peut distinguer 2 phases de 14 jours. La première phase est dite
oestrogénique ou folliculaire. Elle commence le premier jour des règles et correspond à la
maturation finale du follicule qui sécrète les œstrogènes. La deuxième phase est appelée
progestative ou lutéale. Elle correspond au développement du corps jaune qui sécrète à la fois
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
1
/
67
100%