Mécano-sensibilité active des cellules ciliées de l`oreille interne

Rapport de séminaire de la FIP rédigé par Pierre Paga
Mécano-sensibilité active des cellules ciliées
de l'oreille interne
Introduction :
Ce séminaire a été présenté par Pascal Martin, de l'institut Curie. L'objectif des travaux
exposés est de comprendre comment l'oreille parvient à un tel degré de sensibilité dans la détection
des vibrations sonores.
Lorsque nous parlons, nos cordes vocales occasionnent en vibrant une variation locale de
pression. Cette perturbation, qui se propage dans l'espace, est caractérisée par :
●une fréquence
●une intensité (ou amplitude)
Ce stimulus sonore entraine une vibration du tympan, qui se propage à trois osselets : le
marteau, l'enclume et l'étrier. Elle atteint ensuite la cochlée et le vestibule.
Les cellules de la cochlée opèrent alors une conversion du stimulus mécanique en signal
électrique : c'est la transduction. Ce signal atteint ensuite le cerveau par l'intermédiaire du nerf
auditif.
L'oreille est un détecteur remarquable :
●gamme fréquentielle : l'oreille peut détecter des fréquences allant de 20Hz à 20kHz chez
l'homme, et de 20Hz à 100kHz chez les baleines. L'oreille est donc un détecteur rapide, avec
des temps de réaction inférieurs à 10μs.
●Sensibilité : l'oreille peut détecter des sons d'intensité comparable à ceux du bruit
environnant. Cela revient à détecter des vibrations d'amplitude inférieure au nanomètre. De
plus, la discrimination fréquentielle (intervalle de fréquences entre deux signaux perçus
comme distincts) est de l'ordre de 2Hz (
f
f~0.5
)
●gamme d'intensité : la gamme d'intensité perçue s'étend de 20μPa à 20 Pa, soit une gamme
de six ordres de grandeurs. Si l'on considère l'énergie, qui est une fonction quadratique de
l'intensité, celle-ci s'étend sur douze ordres de grandeurs.

Cette gamme immense entraine la nécessité d'une réponse non linéaire de l'oreille. Sinon, les
amplitudes caractéristiques de vibrations s'étaleraient de 1nm à 1mm ! En psychoacoustique, on
utilise donc une échelle logarithmique : l'intensité sonore y est définie par
I=20×logP
Pref
, avec Pref = 20μPa et I en décibels (dB).
Un premier paradoxe apparaît : les organes de l'oreille sont immergés dans un fluide
visqueux. Comment des vibrations d'amplitude très faible peuvent elles se maintenir et être
détectées dans ces conditions ?
Ce paradoxe suggère que l'audition, comme le dit Thomas Gold, est un phénomène actif : il
y a intervention de sources d'énergie non mécaniques.
Une illustration frappante de ce phénomène est que l'oreille est capable d'émettre des sons !
Ce sont les émissions oto-acoustiques, de l'ordre de 0 dB (soit un peu plus faible que le bruit
environnant).
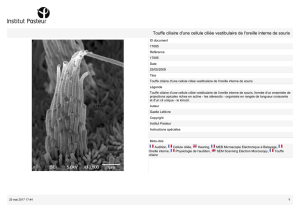
La cellule ciliée
La cellule ciliée est l'acteur final de la perception du son. Une cellule ciliée se présente
comme une cellule surmontée d'un faisceau appelée touffe ciliaire. Ces cellules sont organisées en
rangées :

La touffe ciliaire est composée de stéreocils arrangés de façons régulière. Ce sont des
bâtonnets d'actine (un composant des membranes cellulaires) possédant la propriété d'être plus fins
à la base qu'au sommet :
En conséquence, lorsque, en laboratoire, une force latérale est exercée sur eux, ils cisaillent
à la base. C'est ce mouvement qui crée le signal électrique. Par quel mécanisme ?
Pour visualiser le phénomène, on injecte dans la cellule un marqueur fluorescent sensible au
calcium (les ions Ca2+ sont absents de l'intérieur de la cellule mais abondants a l'extérieur).
Lorsque la touffe ciliaire est mise en mouvement, on voit une illumination de la cellule. Le
déplacement de la touffe ciliaire permet donc au calcium de pénétrer dans la cellule.
En fait, le phénomène est plus complexe. Le calcium pénètre dans la touffe ciliaire, mais est
immédiatement pompé au dehors. Ce n'est pas le cas du potassium (K+), qui parvient à pénétrer
dans la cellule. Or habituellement, la cellule est polarisée électriquement par rapport au milieu
extérieur (la différence de potentiel est de l'ordre de 60mV). L'irruption du calcium provoque donc
une légère dépolarisation de la cellule. Ce qui, à son tour, provoque l'ouverture de canaux
perméable au calcium, qui pénètre alors la cellule, entrainant l'ouverture des canaux d'évacuation du
potassium et la repolarisation de la cellule. Ce cycle entraine l'activation de la synapse adjacente,
qui envoie un signal électrique.
Fonctionnement de la touffe ciliaire :
La touffe ciliaire présente des rangées de cils. Le sommet de chaque cil est connecté au flanc
du cil de la rangée adjacente.

La longueur du fil de liaison est d'environ 200nm. Lorsque la touffe ciliaire est mise en
mouvement, la tension de ces fils varie, ce qui pilote l'ouverture de canaux tensiosensibles (une
tension élevée entraine l'ouverture du canal).
Le système canal-fil se comporte comme un ressort accroché à une porte. L'énergie est
différente selon que la porte est fermée ou ouverte. En effet, l'ouverture de la « porte » réduit la
longueur du ressort d'une longueur d. Il y a donc une différence d'énergie
E
entre l'état ouvert
et l'état fermé du canal. La probabilité d'ouverture du canal est alors donné une fonction de
E
,
donnée par la statistique de Maxwell-Boltzmann.
Les touffes ciliaires sont de plus capables de vibrer en l'absence de stimulus, comme le
montrent des observations réalisées en chambres sourdes. Leur amplitude varie de 25nm à 80nm.
L'analyse de ces oscillations montre un contenu fréquentiel concentré entre 5Hz et 15Hz, avec un
maximum à 8Hz, avec un facteur de qualité correspondant de l'ordre de 1.
Pour sonder ces oscillations, on entraine en mouvement les touffes ciliaires à une fréquence
proche de leur fréquence caractéristique. On utilise pour cela une fibre artificielle flexible, qui
permet de mesurer la force exercée par la touffe. La fréquence d'agitation est de 9Hz.
En l'absence de stimulus, le mouvement est bruité, irrégulier. En présence de stimulus, au
contraire, le mouvement est très régulier, en phase avec le stimulus. De plus, l'amplitude est le
double de l'amplitude du stimulus !

On introduit une fonction
= K
F
, correspondant à la sensibilité du système. La
courbe obtenue présente un maximum de 5 aux alentours de 10 Hz, et tend vers 1 aux hautes
fréquences, alors que la cellule non oscillante de référence se maintient à 1. De plus, le diagramme
de phase associé présente une avance de phase pour les fréquences inférieures à 9Hz, ce qui
caractérise un système actif.
D'après le théorème de fluctuation-dissipation,
×C
2×kT ×' ' =1
pour un système
passif, avec
' '=ℑ
, et
C
le spectre des fluctuations. Or ici, il y a divergence aux
alentours de 9Hz !
On observe aussi que près de la fréquence 8Hz, les petits stimuli sont multipliés par 10. La
touffe ciliaire possède donc les caractéristiques d'un filtre passe bas.
L'étude effectuée avec plusieurs cellules montre que le gain passe d'environ 7 à 1000 lorsque
l'on passe d'une cellule à plusieurs.
A quoi correspondent les oscillations ?
La manipulation suivante est effectuée : on déplace la touffe ciliaire d'une longueur x, et on
mesure la force exercée par la touffe en retour:
Lorsque les déplacements sont grands, la réponse de la touffe est linéaire : elle correspond à
celle d'un ressort dont la constante de raideur est donnée par la pente de la courbe. Mais pour de
faibles déplacements, la touffe ciliaire se comporte comme un ressort de raideur négative !
Le profil énergétique montre deux minima : en -20 nm et en +20 nm. L'oscillation s'effectue par
transit entre ces deux minima.
 6
6
1
/
6
100%