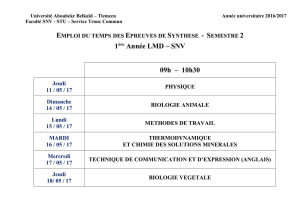
Modèles Impulsionnels de Neurones: Théorie et Applications

http://www.snv.jussieu.fr/brette/
Modèles Impulsionnels de
Neurones: Théorie et Applications
Romain Brette
INSERM U483

http://www.snv.jussieu.fr/brette/
Le neurone
Les neurones communiquent
par impulsions électriques
(potentiels d’action).
10 mV
1 s

http://www.snv.jussieu.fr/brette/
Modèles impulsionnels
Impulsions →impulsions

http://www.snv.jussieu.fr/brette/
Modèles impulsionnels
Variable continue →impulsions
(encodeur)

http://www.snv.jussieu.fr/brette/
Formulation générale
1) Une équation différentielle:
2) Un mécanisme de réinitialisation:
impulsion quand V > seuil
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
1
/
47
100%