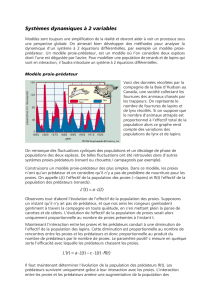
Projet Mathématiques Numériques Évolution d`espèces





 6
7
8
9
10
11
12
13
6
7
8
9
10
11
12
13
1
/
13
100%