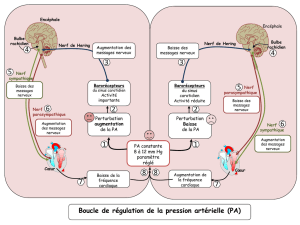
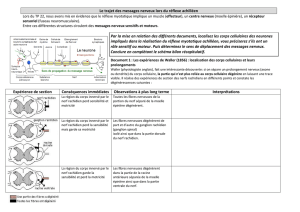
Cours-Prof: Le tissu nerveux

La Moelle
Épinière
Potentiel
D’action
Monophasique

2
²LYCEE TAIBA/I.C.S. DE MBORO
tél : 955 77 81 – 955 40 37 - bp : 35 - site web : www.lmboro.has.it-e-mail : lticsmboro@sentoo.sn
Thème I: ORGANISATION DU SYSTHEME NERVEUX
CEREBRO-SPINAL DES VERTEBRES
* * *
Thème II : LE TISSU NERVEUX : ORGANISATION
ET PROPRIETE
* * *
Thème III : RÔLE DU SYSTHEME NERVEUX DANS
LE COMPORTEMENT MOTEUR D’UN ANIMAL
* * *
Session : 2007 / 2008

3
THEME I- ORGANISATION DU SYSTHEME NERVEUX
CEREBRO-SPINAL DES VERTEBRES
INTRODUCTION
L’étude anatomique permet de diviser le système nerveux en deux parties :
- Les centres nerveux ou système nerveux central ou névraxe constitué de
l’encéphale et de la moelle épinière : on parle de système nerveux
cérébrospinal ;
- Les nerfs ou système nerveux périphérique qui forment un vaste réseau dont
les ramifications atteignent les moindres recoins des tissus de l’organisme
doués d’une sensibilité et qui présentent par endroit des nœuds ou ganglions.
On distingue 12 paires de nerfs crâniens et 33 paires de nerfs rachidiens
Et du point de vue physiologique, le système nerveux se divise en :
- Système nerveux de la vie de relation formé par les centres nerveux d’où
partent les nerfs céphaliques et rachidiens qui innervent les muscles forts de
l’organisme (muscles striés squelettiques) ;
- Système nerveux de la vie de nutrition ou système nerveux végétatif ou
système nerveux sympathique, formé par les portions du névraxe d’où partent
les nerfs qui contrôlent la plupart des fonctions automatiques et inconscientes
de l’organisme. Il est constitué de deux système antagoniques : le système
orthosympathique et le système parasympathique qui gouvernent ainsi les
muscles lisses du corps et qui innervent le coeur.
I- ORGANISATION DE L’ENCEPHALE : EXEMPLE CHEZ LE
MOUTON
I-1- Morphologie externe
I-1-1- Face dorsale (face supérieure ou postérieure)
De l’avant vers l’arrière on observe :
a- le cerveau : formé de deux masses appelées hémisphères cérébrales séparées
par le sillon inter hémisphérique. Celles-ci ont des surfaces qui présentent un
aspect plissé du fait qu’elles sont parcourues de sillons délimitant des
circonvolutions cérébrales.
b- le cervelet : Situé en avant du cerveau, il est formé d’un lobe médian ou vermis
médian qui sépare deux autres lobes appelés hémisphères cérébelleux dont les
circonvolutions sont plus fines que celles du cerveau.
c- Le bulbe rachidien : Il est rattaché en parti au cerveau et se poursuit par la
moelle épinière.
I-1-2- Face ventrale (face inférieure ou antérieure)
L’observation montre qu’elle est formée de l’avant vers l’arrière par :
- Les lobes olfactifs point de départ des nerfs olfactifs ;

4
- Deux nerfs optiques formant un chiasma ou chiasma optique ;
- La cicatrice de l’hypophyse ;
- l’hypothalamus qui relie l’hypophyse à l’encéphale par un pédoncule ;
- Les pédoncules cérébraux qui sont de gros cordons blancs qui s’engagent dans un
anneau transversale ou protubérance annulaire (ou pont de varole) ;
- Le bulbe rachidien qui prolonge les pédoncules cérébraux.
Remarque :
Les pédoncules cérébraux, le pont de varole et le bulbe rachidien forment le tronc
ou axe cérébral d’où se partent les 12 paires de nerfs crâniens.
I-2- Etude de coupes
I-2-1- Coupe longitudinale (par le sillon inter hémisphérique, milieu des couches
optiques, milieu du cerveau)
L’observation d’une coupe longitudinale de l’encéphale de mouton permet de
distinguer cinq principales parties :
a- Le cerveau antérieur ou télencéphale : Il comprend les hémisphères cérébraux
qui cachent des lames blanches, le corps calleux et le trigone, qui les unissent.
Entre les deux lames s’étend une cloison transparente qui sépare la cavité en deux
ventricules : le ventricule latéral droit et le ventricule latéral gauche. Au fond de
chaque ventricule rempli du liquide céphalorachidien, se trouve une masse, le corps
strié.
b- Le cerveau intermédiaire ou diencéphale : Une légère incision en bas du trigone
montre une cavité appelée troisième ventricule. Les parois de ces trois cavités (1er,
2ième et 3ième) sont formées de couches optiques ou thalamus. L’épiphyse est
rattaché au plafond du 3ième ventricule et est relié à l’hypothalamus qui se trouve
sur les planchés. Le 3ième ventricule communique avec les deux ventricules latéraux
par le trou de Monro. Ces deux couches optiques sont reliées par la commissure
grise.
C - Le cerveau moyen ou Mésencéphale : A ce niveau le 3ième ventricule se rétrécit
et forme l’aqueduc de sylvius. Ce dernier est limité dorsalement par les tubercules
quadrijumeaux ou couches optiques et ventralement par les pédoncules cérébraux.
D- Le cerveau postérieur ou Métencéphale :A ce niveau, nous constatons que le
4ième ventricule fait suit à l’aqueduc de Sylvius antérieurement (ventrale) et
postérieurement (dorsale) par le cervelet qui est formé d’une substance grise
externe ou écorce cérébelleux et d’une substance blanche interne ou arbre de vie.
Le cervelet est relié à l’encéphale par trois paires de cordons blancs ou pont de
varole.
E- L’arrière cerveau ou Myélencéphale: Il est formé par le bulbe rachidien qui est
la racine de la moelle épinière et qui présente une cavité ou 4ième ventricule qui se
prolonge par le canal de l’épendyme dans cette dernière (la moelle épinière).
I-2-2- Coupe transversale (au niveau du diencéphale : thalamus chiasma)

5
Celle-ci permet d’observer la position des ventricules et des différentes parties de
l’encéphale qui sont toutes remplies par le liquide céphalorachidien ou liquide
cérébrospinal.
Cette coupe permet de constater aussi que le cerveau est formé d’une substance
grise externe ou matière grise et d’une substance blanche interne : on dit que le
cerveau présente une dualité structurale.
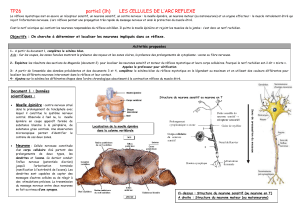
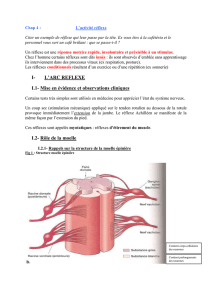
II- ORGANISATION DE LA MOELLE EPINIERE ET DES NERFS
II-1- Morphologie externe :
La moelle épinière est un long cordon blanc nacré logé dans le centre du canal
rachidien, dans la colonne vertébrale et elle est de forme cylindrique.
II-2- Etude de coupe transversale de la moelle épinière (avec racines des
nerfs rachidiens)
Une coupe transversale nous permet d’observer :
a- Un creux ou sillon dorsal ou postérieur étroit et prononcé (profond) au niveau de
sa face dorsale et un creux ou sillon ventral ou antérieur large et peu profond au
niveau de sa face ventrale ;
b- Une dualité structurale contraire à celle de l’encéphale avec :
- Une substance blanche externe formant les cordons postérieurs, latéraux et
antérieurs ;
- Une substance grise interne formant les cornes postérieures, corne
intermédiaire et antérieures.
c- Des excroissances latérales qui correspondent à des racines postérieures
longues, fines et munies chacune d’un ganglion spinal et des racines antérieures plus
court et plus large dépourvues de ganglion spinal. De part et d’autres du canal de
l’épendyme (trou central de la moelle épinière rempli de liquide céphalorachidien),
une racine postérieure et une racine antérieure s’unissent pour former un nerf
rachidien.
REMARQUES
1- les protections du système nerveux : Le système nerveux est protégé par :
a- les os : Ils s’agissent des quatre os qui forment la boite crânienne pour
l’encéphale et des vertèbres (32 à 33) de la colonne vertébrale pour la moelle
épinière ;
b- Les membranes ou méninges :
- La dure-mère : de nature fibreuse, collée aux os, elle est la plus externe et a
un rôle purement protectrice ;
- L’arachnoïde : elle est de nature feuilleté et présente des lacunes remplies
par le liquide céphalorachidien qui présente la même composition chimique que
le plasma ; elle joue donc en même temps un rôle nourricier et de suspenseur
hydraulique (rôle de défense) ;
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
1
/
34
100%