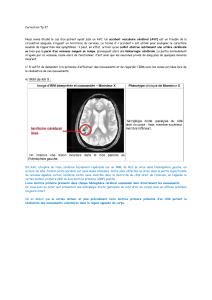
thèse imagerie mentale motrice contrôlée par interface cerveau

UNIVERSITÉ DE NANTES
FACULTÉ DE MÉDECINE
ANNÉE 2010
THÈSE
pour l’obtention du
DIPLÔME D’ÉTAT DE DOCTEUR EN MÉDECINE
(DES DE NEUROCHIRURGIE)
Présentée et soutenue publiquement le
20 Octobre 2010
par
Vincent ROUALDES
Né le 2 août 1976
à Poitiers (Vienne)
IMAGERIE MENTALE MOTRICE CONTRÔLÉE
PAR INTERFACE CERVEAU-MACHINE :
UNE PORTE OUVERTE SUR LE SYSTÈME MOTEUR
DANS LA RÉCUPÉRATION DES DÉFICITS
RATIONNEL
APPLICATION AUX DÉFICITS DU MEMBRE SUPÉRIEUR
APRÈS ACCIDENT VASCULAIRE CÉRÉBRAL
CAS EXPÉRIMENTAL
et
PROPOSITION DE PROTOCOLE DE RECHERCHE CLINIQUE
Jury
Mr. NGUYEN J.P. Professeur des Universités Président, Directeur
Mr. de KERSAINT-GILLY A. Professeur des Universités
Mr. DERKINDEREN P. Professeur des Universités
Mr. GUILLON B. Praticien Hospitalier
Mme STEFAN A. Praticien Hospitalier
Mme LUCAS M.F. Maître de Conférence des Universités
1

SOMMAIRE
1|Récupération des déficits moteurs du membre supérieur après un accident
vasculaire cérébral
Histoire naturelle de la récupération des AVC et de la fonction du membre supérieur
Evaluation de l’effet des thérapeutiques de rééducation sur la récupération des déficits du membre
supérieur après AVC et place de l’imagerie mentale motrice
2|Pratique de l’imagerie mentale motrice et atteinte du système nerveux central
3| Principes de fonctionnement des interfaces cerveau-machine (BCI) - Application à
la réhabilitation des déficits
Signal cérébral utilisé pour l’interface cerveau-machine
Traitement du signal
Apprentissage du contrôle et de l’utilisation d’un système BCI
Principes neurophysiologiques utilisés pour le contrôle du BCI utilisant le signal
électroencéphalographique
Utilisation du BCI pour la restauration de fonction du système nerveux central
Apprentissage du contrôle du signal cérébral pour sa normalisation
Apprentissage du contrôle du signal cérébral pour la commande d’un dispositif robotisé
assistant le mouvement
4| Anatomie fonctionnelle du système moteur cortical : bases fondamentales pour les
applications d’interface cerveau-machine
Planification de l’action
Cortex moteur primaire M1
Aires frontales à fonction prémotrice
Cortex prémoteur dorsal
L’aire motrice supplémentaire
Cortex somesthésique
Cortex pariétal postérieur
5| Principes neurophysiologiques du codage neuronal dans le système moteur pour
l’interface cerveau-machine
Principe du codage distribué
Principe d’insuffisance du neurone individuel
Principe du neurone multitâche
Principe de masse critique neuronale
Principe de dégénérescence du codage neuronal
Principe de plasticité
3

6| Plasticité cérébrale au cours de la récupération des AVC
Facteurs contribuant à la récupération
Connectivité diffuse
Redessinement des régions voisines
Règles d’apprentissage synaptique au cours de la récupération
Plasticité homéostatique
Plasticité Hebbienne
7| Cas clinique expérimental
Présentation générale
Phénomène neurophysiologique utilisé pour l’interface BCI
Acquisition du signal
Système d’interface cerveau-machine (BCI) utilisé
Chronologie expérimentale
Résultats
A propos d’un cas de la littérature
8| Etat des connaissance sur l’utilisation des rythmes sensori-moteurs pour les
opérations de BCI par imagerie mentale motrice. Perspectives pratiques pour la
réhabilitation des déficits
Spectres fréquentiels utilisables pour le contrôle de l’imagerie mentale motrice
Mécanismes mis en jeu dans le blocage de la transmission corticospinale au cours du processus
de mentalisation
Implication du cortex moteur ipsilatéral dans la rééducation
Classification des mouvements
9| Utilisation de l’imagerie fonctionnelle et de la stimulation magnétique
transcrânienne comme outils d’évaluation des effets d’une réhabilitation des déficits
basée sur l’imagerie mentale contrôlée par Interface cerveau-machine
Imagerie par résonance magnétique
IRM morphologique
IRM en Tenseurs de diffusion
IRM fonctionnelle
Analyse fonctionnelle dynamique
Stimulation magnétique transcrânienne
10| Conclusion
11| Proposition de protocole de recherche clinique
4

INTRODUCTION
Les déficits du membre supérieur après accident vasculaire cérébral représentent un véritable
challenge pour les thérapeutes. Six mois après l’accident, 30 à 60% des patients qui ont une
atteinte initiale sévère n’ont récupéré aucune fonction à ce niveau et seulement 5 à 20% auront
récupéré complètement. Or cette fonction joue un rôle clé dans l’autonomie des patients.
L’impact sur la récupération fonctionnelle des interventions de rééducation confronté à
l’histoire naturelle de ces déficits est très discuté dans la littérature, en particulier concernant la
motricité fine distale. Les améliorations s’appuient essentiellement sur des stratégies
compensatrices.
Parmi le large panel d’interventions proposé et évalué, l’imagerie mentale motrice semble
présenter à la lumière de quelques études bien conduites des effets tout à fait significatifs sur
l’amélioration de la fonction du bras. Elle peut être définie comme la représentation consciente
d’une action et se base sur une activation subliminale du système neuronal moteur.
L’entraînement par imagerie mentale est pratiqué chez le sportif de longue date pour
perfectionner ou acquérir de nouvelles habiletés motrices. En rééducation l’objectif de
l’évocation cognitive d’une action physique est d’augmenter les capacités de mouvement
orienté vers un but ou de stabiliser un mouvement donné formaté.
Elle n’est pourtant pas intégrée systématiquement en pratique courante, sans doute parce
qu’elle pose un problème essentiel : son caractère subjectif. Ceci est d’autant plus gênant chez
des patients pour lesquels des atteintes neuropsychologiques peuvent s’associer à des degrés
variables et sont susceptibles de dégrader cette faculté cognitive, plus complexe qu’il n’y paraît.
Les évolutions technologiques récentes nous livrent de nouveaux outils dont l’impact est déjà
majeur dans le domaine des neurosciences. Appelés Interfaces cerveau-machine ou BCI pour
Brain-Computer Interfaces, ces dispositifs permettent d’établir une communication directe entre
cerveau et machine. Ils enregistrent l’activité cérébrale produite par une tâche mentale et la
convertissent en direct en une commande qui peut servir à contrôler n’importe quelle machine.
Bien mieux encore, ils peuvent fournir au sujet un retour instantané qualitatif et quantitatif de
son activité cérébrale. Voici une solution pertinente pour «monitorer» une tâche d’imagerie
mentale motrice et permettre au sujet d’apprendre à la contrôler.
La plasticité cérébrale permet l’apprentissage; il dépend fortement de l’entrainement. Comment
une rééducation basée sur l’entrainement mental moteur dirigé par BCI peut-elle induire une
récupération motrice ?
5

Nous abordons le sujet pour tenter de répondre à cette question sous un angle atypique, requis
par l’abondante littérature s’y rapportant et par l’imbrication inextricable des notions
fondamentales nécessaires à sa compréhension globlale. Les différents chapitres s’enchaineront
donc selon un continuum débutant par la problématique clinique et thérapeutique. Une porte
est ouverte sur le système moteur grâce à l’interface cerveau-machine; ses principes de
fonctionnement et son intérêt pour le guidage de l’imagerie mentale motrice seront abordés
dans un second temps. Des notions fondamentales incontournables sur l’anatomie
fonctionnelle, les principes neurophysiologiques du codage neuronal dans le système moteur et
la plasticité cérébrale au cours de la récupération feront l’objet de trois chapitres, avec un souci
permanent d’interaction entre ces données et leur application concrète : la compréhension du
rationnel d’une rééducation par l’imagerie mentale motrice guidée et contrôlée à l’aide d’un
système BCI. Nous présenterons ensuite un cas clinique expérimental réalisé au sein du pôle de
neurosciences de Nantes, dont les résultats surprenants ont catalysé notre intérêt vis à vis de
cette technologie et nous conduisent à proposer pour finir un protocole de recherche clinique
basé sur cette technologie qui mérite d’être évaluée scientifiquement.
6
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
1
/
125
100%