Imprimer au format PDF

Retour à Physiologie musculaire
Anatomie des muscles et des cellules musculaires
Chacune des extrémités d?un muscle est attachée à un os par des tendons. Le raccourcissement du muscle change
la relation physique entre ses points de fixation et génère donc un mouvement. En général, les muscles
squelettiques ont leurs extrémités attachées à deux os reliés par une articulation ; le raccourcissement, ou
contraction, de ces muscles provoque alors un mouvement de flexion ou d?extension, tout dépendant de la position
des points d?insertion par rapport à l?articulation. La contraction de certains muscles peut aussi entraîner des
mouvements de rotation, comme par exemple les mouvements d?orientation de la tête, et les mouvements de
pronation et supination de l?avant bras. Ainsi, les muscles squelettiques se contractent toujours longitudinalement,
et le mouvement qu?ils engendrent dépend de la position des points d?insertion par rapport aux leviers que sont les
os.
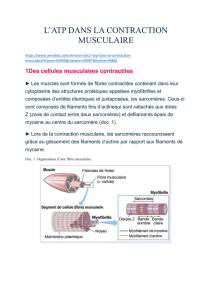
Un muscle squelettique est constitué de plusieurs cellules, ou fibres musculaires organisées en faisceaux parallèles
sur toute la longueur du muscle. À l?intérieur, chaque fibre musculaire est constituée de plusieurs myofibrilles
contractiles parallèles et s?étendant sur toute la longueur de la fibre musculaire. La membrane des cellules
musculaires porte le nom de sarcolemme et son cytoplasme porte le nom de sarcoplasme. Le sarcoplasme de
chaque cellule musculaire comporte un réseau de tubules transverses, ou tubules T, i.e. des invaginations de la
membrane plasmique qui traversent la cellule de part en part vis-à-vis des disques Z. Ces tubules fournissent à la
cellule une voie pour l?acheminement des substances nutritives et de l?oxygène ainsi que pour l?élimination des
déchets. En effet, les cellules musculaires possèdent un gros diamètre, ce qui ralentie les échanges par simple
diffusion. Les tubules augmentent le ratio S/V des cellules et accélèrent ainsi les échanges avec le liquide
extracellulaire. Les tubules T jouent aussi un autre rôle très important : ils conduisent l?influx nerveux, généré à la
surface de la cellule par la liaison de l?acétylcholine, jusque dans les profondeurs de la cellule; ils permettent donc
de diminuer le délai du couplage excitation-contraction.
Toutes les cellules possèdent dans leur cytoplasme un réseau de canaux intracellulaire appelé réticulum
endoplasmique. Le RE joue habituellement un rôle important dans la synthèse des protéines. Dans les cellules
musculaires celui-ci joue un autre rôle important dans le couplage excitation contraction et porte le nom de
réticulum sarcoplasmique. Le RS comporte des structures appelées citernes terminales qui sont des réservoirs
prenant en sandwich les tubules T. Au niveau de la membrane des citernes terminales se trouvent des pompes à
Ca+ utilisées pour séquestrer le Ca2+ dans le RS et ainsi maintenir une faible concentration de Ca+ dans le
sarcoplasme.
Structure des myofibrilles
Chaque myofibrille est un cylindre fait de plusieurs sarcomères, i.e. des sections de cylindre mise bout à bout. Les
sarcomères sont les unités contractiles des cellules musculaires. Chacun est délimité par deux disques Z; entre les
disques Z se retrouvent des filaments contractiles de deux types, qui s?étendent selon l?axe longitudinal du
sarcomère et qui se chevauchent partiellement.
La_contraction_musculaire
Anatomie des muscles et des cellules musculaires 1

Les filaments fins, constitués d?actine, sont attachés aux disques Z, mais libres vers l?intérieur du sarcomère ; les
filaments épais, faits de myosine, sont attachés au centre du sarcomère et non aux extrémités. Dans la région de
recouvrement entre les deux types de filaments, chaque filament de myosine est entouré de 6 filaments d?actine,
chacun d?eux entourés de 3 filaments de myosine. Tout au long d?une myofibrille, des zones où les deux types de
filaments se chevauchent alternent avec des zones sans chevauchement ; c?est cette alternance qui est responsable
de l?allure striée des muscles squelettiques en microscopie. Le mécanisme de la contraction musculaire repose sur
une interaction entre les deux types de filaments entraînant le rapprochement des disques Z de chaque sarcomère et
donc leur raccourcissement.
Structure des filaments contractiles
Les filaments fins d?actine sont des hélices à double brin. Chacun de ces brins torsadés est constitué de monomères
d?actine globulaire. Les filaments fins ressemblent donc à des colliers de perles torsadés. Dans les sillons de
l?hélice constituée par chaque filament d?actine, se retrouve une autre protéine filamenteuse, la tropomyosine ; elle
ressemble à une ficelle qui s?enroule autour du collier de perles torsadé. Environ à tous les 40 nm, la troponine, un
complexe formé de plusieurs protéines globulaires, est attachée à la tropomyosine. À l?état de repos des cellules
musculaires, la troponine et la tropomyosine empêchent qu?il y ait interaction entre les filaments fins et épais (la
tropomyosine bloque l?accès aux sites de fixation de la myosine sur l?actine).
Les filaments épais de myosine possèdent une structure un peu plus complexe. Chaque filament épais est constitué
de plusieurs molécules ressemblant à des bâtons de golf. 2 chaînes longues en double hélice forment la queue
(manche) et le cou de ces molécules. Chacune de ces molécules comporte aussi une double tête globulaire faite de
différentes chaînes protéiques. Un filament de myosine est fait de plusieurs de ces molécules agencées selon un
patron précis. Les queues des molécules sont orientées vers le centre du filament et les têtes font saillies et sont
orientées de façon opposée sur chaque demi-filament. Les têtes constituent des ponts moléculaires qui interagiront
avec les filaments d?actine. Les ponts se répètent sur toute la longueur du filament épais par groupes de 3 disposés
autour du filament et séparés par un angle de 120°. Comme les queues sont orientées vers le centre, celui-ci (zone
H) ne contient pas de têtes de myosine.
La_contraction_musculaire
Structure des myofibrilles 2

Le rôle du calcium dans le couplage excitation-contraction
Le mécanisme de la contraction musculaire repose sur une interaction entre les filaments fins et épais des
sarcomères. Pour générer une tension dans un muscle, les têtes, ou pont de myosine, s?accrochent aux filaments
d?actine et les font glisser vers le centre du sarcomère, les raccourcissant ainsi.
La contraction d?un muscle est déclenchée par un neurotransmetteur libéré par un neurone moteur. Le
neurotransmetteur se lie aux récepteurs des fibres musculaires et y déclenche un influx nerveux (suite à un PPSE).
L?influx nerveux produit par la membrane de la cellule musculaire est propagé par les tubules transverses (tubules
T) et traverse rapidement toute l?épaisseur de la fibre musculaire. Le passage de l?influx nerveux entraîne
l?ouverture des canaux à Ca2+ du réticulum endoplasmique (réticulum sarcoplasmique), ce qui cause l?entrée de
Ca2+ dans le cytoplasme (sarcoplasme). Le calcium en concentration élevée dans le cytoplasme se lie massivement
aux molécules de troponine, qui changent de conformation et déplacent la tropomyosine; ce déplacement permet
l?établissement de liens entre les têtes de myosine et les filaments d?actine. La liaison d?une tête de myosine
entraîne sa rotation, ce qui génère une tension dans les queues des molécules de myosine. Cette tension est
responsable du glissement des filaments fins vers le centre du sarcomère et donc de son raccourcissement.
Voir l'animation
On peut comparer cette interaction à une compétition de souque à la corde. Au signal (calcium), toute l?équipe
agrippe la corde et tire, générant une tension qui entraîne l?équipe adverse vers le centre.
Une autre analogie intéressante est celle du traineau tiré par un chien : le chien tire le traineau à l'aide d'un
élastique. Lorsqu'il avance, l'élastique s'étire et le traineau est tiré lorsque l'élastique reprend sa forme.
Voir l'animation
La_contraction_musculaire
Structure des filaments contractiles 3

Le rôle de l?ATP dans le déroulement de la contraction
L?ATP entre en jeu à ce moment dans le mécanisme de la contraction. En effet, les ponts de myosine doivent
pouvoir se fixer et se détacher de l?actine de manière cyclique pour que le glissement se produise. En absence
d?ATP la liaison actine-myosine est stable ; c?est la rigidité cadavérique. Mais lorsque l?ATP se lie au complexe,
la tête de myosine change de conformation entraînant la séparation du complexe en myosine-ATP et actine. La tête
de myosine étant dotée d?une enzyme ATPase, elle hydrolyse l?ATP, mais le produit de l?hydrolyse, ADP + Pi,
demeure attaché à la tête. La liaison de l?actine au complexe myosine-ADP-Pi entraîne la libération d?ADP + Pi.
Comme la réaction est exergonique (augmentation d?entropie), elle est favorisée par la thermodynamique :
Actine + myosine-ADP-Pi --> Actine-myosine + ADP + Pi
En résumé, la contraction se poursuit tant que le calcium est présent pour rendre les sites de liaisons de la myosine
sur l?actine disponible ; la contraction cesse lorsque l?effet du neurotransmetteur ne se fait plus sentir et que le
calcium retourne dans les citernes terminales du réticulum sous l?action de pompes à calcium. Ces pompes ajoutent
à la consommation d?ATP nécessaire à la contraction musculaire. Pour que la contraction se fasse, il faut aussi que
l?ATP soit présent pour permettre aux têtes de myosine de se détacher pour se rattacher plus loin. Si l?ATP
manque, les filaments demeurent attacher (contractures).
Le contrôle nerveux de la contraction
On appelle unité motrice l?ensemble constitué d?un neurone moteur et de toutes les fibres musculaires qui y sont
reliées. Chaque fois qu?un neurone moteur produit un influx nerveux, toutes les fibres qu?il innerve se contractent.
Les unités motrices peuvent être plus ou moins grandes, i.e. contenir plus ou moins de cellules musculaires pour un
neurone. Par exemple, les unités motrices des muscles des doigts sont plus petites que celle des biceps (plus grand
ratio d?innervation).
Un seul influx nerveux qui se propage sur la membrane d?une fibre musculaire peut normalement produire une
contraction. Il produira une secousse musculaire. Les composantes élastiques du muscle (membrane, tissu
conjonctif) n?ont pas le temps d?être étirées et de développer leur pleine tension. Une secousse isolée ne peut pas
développer la tension maximale dont sont capables les éléments contractiles. Lorsqu?il y a succession d?influx
nerveux rapprochés, la tension musculaire continue d?augmenter jusqu?à l?étirement ou la compression maximale
des composantes élastiques. Lorsque ce niveau d?étirement est atteint, les têtes de myosine continuent à se fixer et
à générer une tension, mais sans raccourcissement des sarcomères. Il s?agit de ce qu?on appelle une contraction
tétanique.
La_contraction_musculaire
Le rôle de l?ATP dans le déroulement de la contraction 4

Imaginez que vous tirez un traineau sur la neige à l?aide d?un élastique. Vous, tirez sur l?élastique, le traineau ne
bouge pas, la tension s?accumule et finalement le traineau avance d?un coup; c?est une secousse musculaire.
Imaginez maintenant que le traineau est attaché à un poteau par un autre élastique; la contraction tétanique
correspond au moment où vous avez complètement étiré l?élastique reliant le traîneau au poteau mais où vous
continuez à tirer en changeant alternativement de main. La tension dans ce deuxième élastique s?ajoute à celle que
vous engendrez dans le premier et entraîne une tension maximale sur le poteau.
L?augmentation graduelle de la tension produite par un muscle repose sur trois mécanismes : 1) le recrutement
d?un plus grand nombre d?unités motrices de plus en plus grosses; 2) la fréquence de stimulation des unités
motrices (+ ou ? de fibres en tétanos) et 3) le type de fibres recrutées (certaines fibres sont plus fortes et plus
puissantes que d?autres).
Mécanique de la contraction musculaire
Relation tension-étirement initial
La tension que produit une fibre musculaire, et par extension un muscle entier, augmente en fonction de
l?étirement initial de celui-ci, jusqu?à un plateau maximal, et diminue ensuite. Ce phénomène s?explique par le fait
que la tension générée par un muscle est proportionnelle au nombre de ponts qui s?établissent entre les deux types
de filaments contractiles ; le nombre de ponts, lui, dépend du degré de chevauchement entre les filaments. Les têtes
de myosine sont réparties régulièrement sur les filaments épais, sauf dans une zone nue au centre appelée zone H.
Lorsqu?un sarcomère est étiré au maximum, aucun chevauchement ne survient entre les filaments, aucune liaison
ne peut s?établir et donc la tension est égale à 0. Lorsque l?étirement est diminué, le nombre de ponts augmente et
la tension aussi, jusqu?à ce que les extrémités des filaments fins effleurent le début de la zone nue ; le
chevauchement est alors maximal, toutes les têtes de myosine des filaments épais peuvent ce lier aux filaments
d?actine et la tension est maximale. Comme un raccourcissement supplémentaire du sarcomères fait pénétrer les
filaments fins dans la zone nue (H), aucun pont ne s?ajoute et la tension demeure inchangée. Lorsque le sarcomère
est raccourci encore plus, les filaments fins commencent à chevaucher les filaments épais de l?autre moitié du
sarcomère ; les liaisons des têtes de myosine sont entravées par ce chevauchement des filaments fins ; les ponts
peuvent alors se fixer aux mauvais filaments fins et produire une force négative ; le nombre de ponts fixés
correctement diminue et la tension aussi diminue.
Le plateau maximal de tension survient à un étirement qui correspond à environ 20 % de l?étirement normal d?un
muscle flasque. Autrement dit, le tonus maintient le muscle dans cet étirement optimal.
Relation force-vitesse
Un tracé de la relation entre la force générée par un muscle et sa vitesse de contraction donne une hyperbole : la
tension produite par un muscle diminue lorsqu?il se contracte plus rapidement pour atteindre 0 à la vitesse
maximale de contraction (Vmax).
La_contraction_musculaire
Le contrôle nerveux de la contraction 5
 6
7
6
7
1
/
7
100%








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}