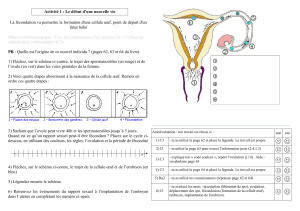
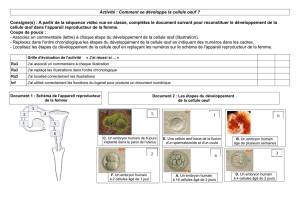
Œufs des Euthériens

Biologie animale : embryologie
Introduction
I. Différentesétapes du développement
Au cours de son cycle de vie tout être vivants subit une reproduction permettant la pérennité
de son espèce. Il existe différent type de reproduction :
- soit par reproduction binaire = simple division de l’organisme mère
- soit par parthénogenèse = un organisme produit un ovule a 2n chromosomes
- soit par reproduction sexué qui fait intervenir des gamètes haploïdes qui refermentdes
gènes parentaux avec une diversité génétique.
Au cours de la gamétogenèse il y a une répartitionaléatoire des chromosomes ce qui entraîne
une diversité génétique.
Les gamètes femellespossèdent des ARNm accumulés lors de la formation ou reçus de cellule
folliculeuse.
Cellule folliculeuse
Ovule
Les ARN sont indispensables pour déclencher le développementultérieur de l’œuf (gamète
male + gamète femelle).
La fécondation peut être :
- externe = aléatoire avec beaucoup de perte
EX : les oursins, les éponges, les poissons…
- interne = le mâle introduit ces gamètes directement dans la femelle
EX : les oiseaux, les mammifères…
- traumatique = le male pique la femelle et introduit ses gamètes dans le sang de la
femelle.
EX : les scorpions, les punaises…
Cloaque = pont commun entre vessie, intestin et appareil reproducteur.
Spermatophore = «sac » ou sont déposées les spermatozoïdes par le male.
Après la fécondation s’enchaînent des phases de développementembryonnaires.
A. La segmentation
Division

Œuf (2n) 2 blastomères (n)
Toutes les divisions issues de la segmentation sont effectuées grâce à l’accumulation des
ARN maternels et continuent par la suite grâce aux maternels
La fin de la segmentation est le stade blastule de l’œuf = blastula
La blastula possèdent un certain nombre de blastomères selon l’espèceétudiée, cette quantité
de blastomèresdépend de la quantité de matière accumulée par l’ovule, d’où une classification
des œufs selon leur matière accumulée (ARN + vitellus (matériel de réserve de l’œuf)) :
- œuf Télolécithe = œuf chargé de réserve
Ex : les œufs de poule.
- œuf Héterolécithe = œuf avec matière de réservedisséminé dans le cytoplasme. L’œuf
n’est pas homogène = œuf Anisotrope.
Ex : les œufs d’amphibiens.
ARN maternel
Vitellus
- œuf Oligolécithe = œuf très peu chargé en vitellus.
Ex : les œufs d’oursins.
-œuf Alécithe = œuf qui n’a pas de matière de réserves,nécessite un apport de réserves
externes pour son développement.
Ex : les œufs desmammifèressupérieurs.
-œuf centrolécithe = œuf chargé en vitellus mais vitellus placé au centre de l’œuf.
Cytoplasme
Vitellus
Ex : les œufs d’insecte
La segmentation varie en fonction de la quantité de vitellus.
Elle peut être totale c'est-à-dire que tout l’œuf se divise
Elle peut aussi être totale et égale, c'est-à-dire qu’il y a la même quantité de matière dans
chaque blastomère nouvellement créé.
[Schéma coupe pole animal pole végétatifPG]
La segmentation peut aussi être totale etinégale =2types de blastomères :
- petit = micromère
- gros = macromère

Les axes de divisions peuvent être horizontaux, verticaux ou inclinés de plus ou moins 30° ce
qui donne une disposition en zigzag des blastomères qui ne permet plus un passage direct
entre le pole animal (PA) et le pole végétatif (PV)
Ex : les Annélides, les Mollusques = division en spirale
Animaux dit SPIRALIA
[Schéma spiraliaPG]
Remarque : le blastomère D est particulier car dans la 4eme division les cellules sœur auront
une destiné particulière pour former la lignée germinale
Si la segmentation est de type bilatéral les blastomères se rangeront immédiatement suivant
les deux axes de l’embryon au fur et à mesure de leur formation.
Axe antéropostérieur
Axe dorso-ventral
Mais la segmentation peut être de type rotationnel :
Compaction
Compaction = cellules très jointives montrant des point de contact. Les cellules s’orientent et
donnent les cellules internes et externes. Au stade 16 les cellules sont orientées et l’organisme
aura sont orientation définitive.
La segmentation holoblastiqueest une segmentation avec compaction.
La segmentation peut êtreméroblastique.Tout le zygote n’est pas concerné par la segmentation
car le vitellus n’est jamais coupé.
Ex : les œufs Télolécithes et centolécithes
Blastule (périblastule)
Syncytium = une cellule a plusieurs noyaux
B. La gastrulation
Dans cette étapeles blastomères vont s’arranger de façon harmonieuse en 3 feuillets
embryonnaires :
- ectoderme (ou ectoblaste)
- endoderme
- mésoderme
Il y a une grande série de mouvements morphogénétiques pour la mise en place des feuillets :
L’épibolie = mise en place par petit mouvement
L’embolie = mise en place par invagination
La délimitation = où des cellules se détachent de l’ensemble et pénètrentdans la blastula pour
former le mésoderme.

C. L’organogenèse
C’est la mise en place des organes et des tissus de l’animal. Chaque type de feuillet donne
un certain organe spécifique :
- ectoderme = épiderme et système nerveux
- endoderme = tube digestif et ses dérivés
- mésoderme = squelette, tissusmésenchymateux,cœur, muscles
Ces mises en place s’effectuent sous l’action de facteurs inducteurs (c’est l’induction).
Ceci nécessite deux intervenants :
- le déterminant
- des cellules compétentes qui réagissent car elles possèdent un récepteurspécifique du
facteur.
Cellule compétente
Mésoblaste
Tube nerveux
L’induction ne se manifeste que pendant un laps de temps déterminé. Ce qui induit que les
conséquences de l’induction ne sont perceptibles que quelques temps après la réception du
signal.

Chapitre 1 : Embryologie de l’oursin
L’œuf est une cellule totipotente = c’est une cellule qui a toutes les possibilités possibles
(beaucoup de potentialité de cellule). Cette totipotence est perdue au cours du
développementembryonnaire.
Ex : blastomèrelividus
C’est un animal gonochorique (sexe séparé) et la fécondation se fait dans le milieu extérieur.
I. Embryologie descriptive
A. L’œuf
L’œuf est relativement petit (≈ 100μm de diamètre) et on observe une bande orangé
subéquatorial au niveau de l’hémisphèrevégétatif.
PA
PV
La charge en vitellus de l’œuf est faible mais ce vitellus est plus abondant dans
l’hémisphèrevégétatif. Comme tout les œufs, l’œuf de l’oursin est anisotrope (pas uniforme).
B. La segmentation
La segmentation est complète et totale = de type holoblastique.
▪ Lapremière division s’effectue une heure après la fécondation, elle estméridienneet donne
naissance aux deux premiers blastomères
Œuf Stade 2
▪ La deuxième division est méridienne mais perpendiculaire à la première.
Stade 4
▪ La troisième division est horizontale et coupe chaque blastomère
Zone animale x4 blastomères
Zone végétative x4 blastomères
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
1
/
23
100%