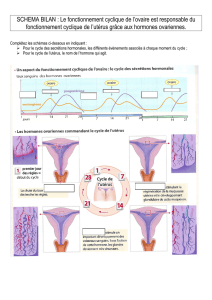
les messagers intracellulaires de l `action hormonale - iPubli

iscic 1987 ; 3: 566-7
L

4,
U
s 8 7
A
(
1
/
2
100%