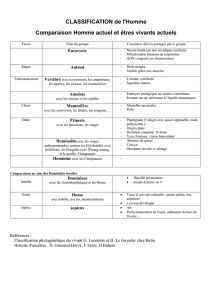
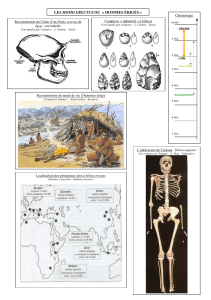
histoire des idees sur l`evolution de l`homme

Thèse présentée pour obtenir le grade de
Docteur de l'Université Louis Pasteur
Strasbourg I
Discipline : Sciences de la Vie
Par Claude STOLL
HISTOIRE DES IDEES SUR
L'EVOLUTION DE L'HOMME
Tome I
Soutenue publiquement le 24.10.2008
Membres du Jury
Directeur de Thèse : M. Jean-Marie VETTER, Professeur, ULP
Co-Directeur de Thèse : M. Jean-Claude GALL, Professeur, ULP
Rapporteur Interne : M. Jean-Jacques ROEDER, DR, CNRS
Rapporteur Externe : Mme Brigitte SENUT, Professeur, MNHN
Rapporteur Externe : M. Josué FEINGOLD, DR INSERM, U.155

2
REMERCIEMENTS
Commencer à rédiger les Remerciements d'une thèse est un moment particulièrement
apprécié, tout d'abord parce que, comme il s'agit de la partie que l'on rédige en dernier, cela
signifie que la rédaction du manuscrit est (enfin !) terminée, et ensuite parce que c'est le
moment où l'on peut faire apparaître la dimension vraie du travail. Une image relativement
courante décrit le thésard seul dans ses questionnements ; en ce qui concerne ce travail, cela
n'a pas été le cas. Au contraire, cette période de réalisation de thèse a été pour nous l'occasion
d'une véritable aventure humaine.
Si l'on suit l'évolution de ce travail, nos remerciements vont au Professeur Jean-Claude
Gall qui nous a accueilli à l'Institut de Géologie et nous a donné les moyens de faire ce travail.
Il a dirigé notre thèse. Il a eu la patience de nous encadrer durant ces cinq années. Nous
voudrions lui exprimer notre reconnaissance pour ses précieux conseils lors de nos
conversation scientifiques, pour son encadrement, sa vision scientifique, sa pédagogie et les
conseils avisés qu'il nous a prodigués tout au long de ce travail de thèse. Nous avons une
lourde dette de gratitude envers lui.
Nous voudrions également exprimer notre gratitude au Professeur Jean-Marie Vetter
passionné d'histoire de la médecine, mais aussi, comme il l'a démontré ici, d'histoire de
l'homme. Il n'a pas compté son temps pour répondre à nos questions et nous conseiller. Ses
grandes qualités d'écoute et son dynamisme ont été particulièrement importants. Sans son
aide, les choses auraient sûrement été bien différentes.
Nous remercions aussi beaucoup la "découvreuse" du plus célèbre fossile des premiers
hommes, Brigitte Sénut, qui nous a si gentiment accueilli et guidé dans notre travail. Ses
précieux conseils nous ont permis d'avancer sur le terrain de la paléontologie, discipline que
nous connaissions très mal. Les articles et les ouvrages que non seulement elle nous a
indiqués (mais aussi donnés ou prêtés) nous ont permis de marcher dans la bonne direction et
de parcourir beaucoup plus sûrement les étapes de notre cheminement.
Nous remercions notre collègue et ami le Docteur Josué Feingold avec qui nous avons
parcouru la plupart des nombreuses années de notre carrière. Il nous a initié avec sa rigueur,
son sens aigu de la pédagogie reconnu et apprécié de tous, à la génétique des populations,
discipline passionnante que certains des acteurs de la théorie synthétique de l'évolution ont
marqué de leur empreinte. Nous le remercions pour tout ce qu'il nous a appris et pour avoir
accepté de juger notre travail.
Nous tenons aussi à remercie le Docteur Jean-Jacques Roeder d'avoir accepté de juger
notre thèse, en prenant du temps sur ses très prenantes recherche en primatologie.
Nous voudrions exprimer notre profonde gratitude au Docteur André Pichot,
certainement le plus grand spécialiste de Lamarck, qui nous a accueilli au tout début de notre
projet et qui nous a fait profiter de sa grande expérience, aussi bien dans les aspects théoriques
que pratiques de notre travail.
Nous remercions aussi les bibliothécaires des bibliothèques strasbourgeoises et
parisiennes que nous avons mis à contribution pour consulter les ouvrages anciens, en
particulier, dont nous avons eu besoin. Leur disponibilité et leur compétence ne nous ont
jamais fait défaut.
Madame Marie-Hélène Chevalier, qui fut jadis, dans notre service hospitalier, une
secrétaire de qualité exceptionnelle, a assuré avec talent, dévouement et patience la
dactylographie et la présentation de notre texte.

3
RESUME
Si l’on ne peut toujours pas répondre avec précision à la question d’où vient l’homme,
on dispose aujourd'hui d'un certain nombre de données qui ont permis de progresser dans
notre compréhension des conditions qui ont conduit à l'émergence de l'humanité à partir des
primates.
Jusqu'au 18ème siècle, la place singulière occupée par l'homme au sein du monde vivant
faisait l'objet de débats entre théologiens et philosophes. La transformation des espèces ne
pouvait se concevoir car la vision du monde qui prévalait alors en Occident reposait sur deux
affirmations majeures: 1) l'univers avait été construit par un Créateur intelligent ; 2) le monde
était statique, invariable. Un changement dans les idées s'opéra progressivement grâce aux
progrès enregistrés dans différents domaines scientifiques : la philosophie naturelle, la
biologie, et la géologie. La période entre 1740 et 1840 fut cruciale pour l'histoire naturelle. Le
concept d'évolution des espèces vivantes était déjà "dans l'air" durant la seconde moitié du
18ème siècle. Certains historiens des sciences n'ont pas hésité à qualifié d'évolutionnistes des
esprits éclairés comme Diderot, Rötig, Herder, Goethe ou Kant. En réalité, ces « novateurs »
étaient des essentialistes qui expliquaient l'apparition de nouvelles formes de vie par des
créations successives. Toutefois, selon Maupertuis (1698-1759) il y avait beaucoup trop de
diversité et d'hétérogénéité dans la nature pour que le monde ait été créé par « dessein ». Il
expliquait l’origine du monde vivant par la génération spontanée d’animaux et de plantes et
par l’élimination des déficients. Cependant, ce dernier mécanisme n’avait rien à voir avec la
sélection naturelle.
Du fixisme au transformisme : l'homme et le singe s'inscrivent dans une histoire :
Le naturaliste Georges-Louis Leclerc de Buffon (1707-1788) fut l'un des premiers à
avoir eu l'intuition de l'immense durée des temps géologiques. En contradiction avec les
affirmations des textes de la Genèse figurant dans l’Ancien Testament, il propose un âge de
75 000 ans pour la Terre au lieu des 6 000 ans de la Bible (il envisagera même des durées de
l'ordre de 3 millions d’années). En outre, il s'intéresse à la variabilité des caractères au sein
des espèces vivantes. Elargissant ses observations, il admet que les individus s’éloignent du
type originel ou idéal en fonction des différences de nature de leur environnement. Buffon
croyait à la possibilité d’une descendance des espèces à partir d’ancêtres communs. L'espèce
n'est plus considérée comme une entité fixe. Désormais les deux obstacles qui s'opposaient à
une conception évolutionniste de la vie étaient levés. La durée considérable des temps
géologiques et la variabilité naturelle des espèces rendaient concevables la transformation des
êtres vivants.
Elève de Buffon, Jean-Baptiste de Lamarck (1744-1829) énonce, dès 1800, les principes
de sa théorie évolutionniste. D'après lui, l’ordre naturel des êtres se caractérise par la
coexistence d'espèces primitives et d'espèces évoluées. Celui-ci résulte de la création continue
des êtres les plus simples (par génération spontanée) et de leur transformation progressive.
Les circonstances, c'est-à-dire l’environnement, sont responsables de la diversité des espèces
car à chaque changement, elles suscitent de nouveaux besoins. En d'autres termes, la fonction
crée l'organe. Bien que Lamarck ne l'ait jamais formulé ainsi, de tels processus impliquent
l’hérédité des caractères acquis. Par rapport à Linné (1707-1778) qui classa l’homme près des
singes, Lamarck va plus loin en faisant descendre l’homme du singe. Dans son ouvrage de
référence "Philosophie zoologique" (1809), il évacue les sciences naturelles du domaine de la
théologie. Lamarck fut critiqué par son contemporain Georges Cuvier (1769-1832),
créationniste notoire, qui récusait l'idée de transformation des espèces. Cuvier reconnaissait
dans l’histoire de la vie l’existence de différentes périodes séparées par des catastrophes qui, à
chaque fois, assuraient le renouvellement des faunes et des flores.

4
Etienne Geoffroy Saint-Hilaire (1772-1844), spécialiste de l’anatomie comparée,
apporte une contribution majeure aux idées de Lamarck. A l'âge de 23 ans, il énonce le
principe de l’unité de plan d’organisation des êtres vivants. La loi des connexions préside
comme cause à l’ordre et à l’harmonie qui règnent dans le monde vivant, tandis que la loi de
balancement rend compte de la compensation des caractères anatomiques. Dès 1833, il
explique, dans un essai, pourquoi des animaux peuvent être différents malgré un plan
d’organisation commun. Geoffroy Saint-Hilaire a eu le mérite d'envisager, pour la première
fois, des liens de descendance entre les espèces disparues et les espèces actuelles.
Le darwinisme : l'homme et le singe ont même origine.
L'année 1859 marque une date-clé dans l’histoire des idées sur l’évolution des espèces.
C’est celle de la publication par Charles Darwin (1809-1882) de : "De l’Origine des espèces
au moyen de la sélection naturelle". Darwin propose une théorie évolutionniste matérialiste
qui fait intervenir la variation et la sélection. Ses idées sont débarrassées de tout préjugé
théologique ou philosophique. La sélection naturelle résulte du fait qu’à chaque génération, il
naît plus d’individus que ne peuvent en supporter les ressources de l’environnement, une idée
empruntée au pasteur et économiste Thomas Malthus (1766-1834). Comment fonctionne-t-
elle? Les individus d’une même espèce sont différents entre eux. Ils sont pourvus de
caractères distincts qui en avantagent certains ou en défavorisent d’autres, selon les conditions
du milieu et selon les partenaires sexuels. Ceux qui survivent sont les plus aptes qui vont se
reproduire et transmettre leurs caractères à la génération suivante. Selon Darwin, qui ignorait
encore les processus de l'hérédité, la micro-évolution suffit à expliquer l'apparition de
nouvelles espèces, voire de nouvelles lignées, ce qui correspond, en fait, à la macro-évolution.
Un tel schéma implique des transformations progressives, plus ou moins régulières. C'est le
gradualisme. Pour Darwin il n'existe pas de changements rapides ou de sauts au cours de
l'évolution. En 1871, Darwin situera l'origine de l'homme dans un pays chaud, à l'Eocène en
s'écartant du groupe des singes catarrhiniens.
Darwin n’est pas seul à avoir proposé que les espèces évoluent sous l’influence de la
sélection naturelle. Avant lui, Alfred Russel Wallace (1823-1913) avait fait une proposition
analogue. Wallace publia en 1855, antérieurement à « De l’Origine des espèces », un article
célèbre "Sur la loi qui a présidé à l’introduction de nouvelles espèces", publication qui est une
préfiguration de la théorie de l’évolution. Wallace qui était convaincu de la réalité de
l’évolution dès 1845, rechercha alors les facteurs responsables du changement évolutif. Il
conclut que ce dernier est dû à la lutte pour l’existence qu'il qualifia de sélection
"stabilisante". Il exposa sa thèse très clairement dans un article de 1858. Bien que Wallace
parvint avant Darwin aux conclusions exposées dans « De l’Origine des espèces », son article
fut lu devant la Linnean Society de Londres, après celui de Darwin, le 1er juillet 1858.
Wallace n'en tint pas rigueur à Darwin, dont il défendit la théorie pour laquelle il proposa le
terme darwinisme.
Un autre défenseur des idées de Darwin fut le biologiste Ernst Haeckel (1834-1919)
dont les travaux exercèrent une immense influence dans le monde scientifique, non seulement
en Allemagne où il était plus connu que Darwin, mais dans toute l'Europe. Son ouvrage le
plus célèbre, "Anthropogénie", fut traduit dans de nombreuses langues. Haeckel situe l'origine
de l'homme à l'Eocène, dans un continent aujourd'hui disparu qu'il appelle la Lémurie, et fait
du Pithécanthrope, découvert par Dubois à Java en 1891, une forme de passage entre les
grands singes et l'homme, le chaînon manquant. Sa contribution majeure à l'histoire de
l'origine des espèces, a été la théorie de la récapitulation : l'ontogenèse récapitule la
phylogenèse c'est-à-dire le développement d'un individu retrace toute l'histoire de sa lignée.

5
Avec Auguste Weissmann (1834-1914), l'essor de la toute jeune science de l'hérédité
apporte des arguments nouveaux à la théorie de l'évolution. Désormais, on élimine
définitivement l’idée fausse de la transmission des caractères acquis en distinguant les cellules
somatiques des cellules germinales qui, seules, transmettent les gènes au fil des générations.
Le mutationnisme : une explication des changements évolutifs
La redécouverte des lois de Mendel par Hugo De Vries (1848-1935) et les généticiens
du 20ème siècle, marque un tournant dans la compréhension des mécanismes de la spéciation.
Avec Thomas Morgan (1866-1945) ils établissent que la variabilité au sein des populations
résulte de mutations. Si les mutations s'avèrent le plus souvent délétères, certaines peuvent
cependant donner naissance à des caractères nouveaux, originaux, quelquefois monstrueux.
La théorie synthétique : l'élaboration d'une vision cohérente de l'évolution.
Dans les années 1940, avec Dobzhansky (1900-1975), Huxley (1887-1975), Simpson
(1902-1984) Mayr (1904-2005), Fisher (1890-1962), Haldane (1892-1964) et d'autres,
s'impose la théorie synthétique de l’évolution encore appelée néodarwinisme. Elle incorpore
les données nouvelles de la génétique et les observations des paléontologues. Elle stipule que
l'évolution s'effectue d'une manière graduelle, la sélection naturelle agissant à la manière d'un
filtre sur la variabilité inhérente à toute population d'êtres vivants. Ses processus entraînent,
au cours du temps, une complexité progressive des organismes.
La théorie neutraliste : le rôle de la dérive génétique.
Chez les primates et chez l’homme, des régions étendues du génome semblent
"silencieuses" : elles se dupliquent et ne sont pas soumises à la sélection ; elles se révèlent
neutres. A partir de ce constat, le biologiste japonais Motoo Kimura (1924-1994) a proposé,
dans les années 1970, une théorie dite neutraliste de l'évolution. Il explique l’évolution des
fréquences des gènes par les seuls effets de la dérive génétique, par un effet
d’échantillonnage, et par des variantes apparues par mutation, et ceci, en l’absence de
sélection. Le neutralisme explique l’essentiel de l’évolution moléculaire et l’extraordinaire
polymorphisme qui caractérise les populations sexuées.
"Evo-dévo" et la génétique du développement : une approche pluridisciplinaire.
1977 va marquer le retour des études évolutionnistes sur l’ontogenèse. On associe, dans
une même approche appelée "évo-dévo", les avancées récentes dans les domaines de la
génétique du développement, de l'embryologie, de l'anatomie et de la paléontologie. Un même
complexe de gènes rend compte de la grande unité reconnue dans les plans d'organisation du
règne animal. L'accent est mis sur des modalités évolutives comme la cooptation, l'exaptation,
les paramorphismes ou les hétérochronies du développement dont la néoténie est un cas
particulier. Les mécanismes génétiques sont progressivement élucidés. Des hétérochronies
survenues dans le développement du cerveau, dans l'ossification du crâne et de la face, dans
la répartition de la pilosité rendent compte de la divergence entre pongidés et hominidés.
L'émergence de l'homme d'après les données de la paléontologie.
Au cours de la deuxième moitié du 19ème siècle et jusqu'au début du 20ème siècle,
prévalut chez certains scientifiques la thèse de l'évolution polyphylétique des primates :
l'homme n’était pas classé à part des singes mais plutôt entremêlé à eux. Au fur et à mesure
que s'accumulèrent les découvertes d'hommes fossiles de plus en plus anciens, l'histoire
paléontologique des hominidés se précisa. Depuis la fin de l'ère tertiaire, il y a 7 ou 8 millions
d'années, se sont succédés les Australopithèques, Homo habilis, Homo erectus, les
Néanderthaliens et Homo sapiens. En règle générale, l'évolution de la lignée humaine est
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
1
/
484
100%