Le lymphocyte est une cellule endocrine - iPubli

SYNTHÈSE
Élisabeth Modigliani
Liliane Gattegno
Le lymphocyte est
une cellule endocrine
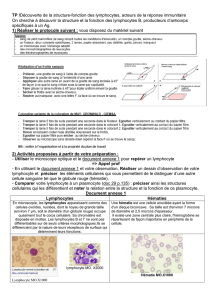
Les lymphocytes peuvent, après activation, synthétiser un
grand nombre de récepteurs d'hormones et de neuromédia
teurs qui modulent leur prolifération et leur fonction ; ils
peuvent aussi produire eux-mêmes certaines de ces hormo
nes qui pourront agir sur le système nerveux central ou sur
des cibles périphériques.
D
à
à
à



b
•
c

;\�
28 1. -
 6
7
6
7
1
/
7
100%