D1-UE10-Diotel-Système_Neurosensoriel_Audition_Equilibre

Page 1 sur 40
UE 10 : SYSTEME NEUROSENSORIEL
N.Diotel
Date : 10/02/2017 Plage horaire : 8h30 – 10h30
Ronéistes : AUPERIN Vanille Enseignant : N.Diotel
BENARD Anne-Lyne
L’Audition et l’équilibre du système vestibulaire
I. Introduction
1) L’audition et la fonction de l’audition
2) Les ondes sonores
3) Les caractéristiques du son
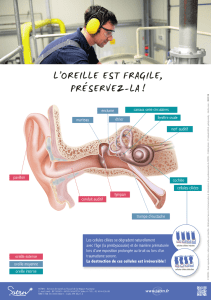
II. L’anatomie de l’oreille
1) L’oreille externe
2) L’oreille moyenne
3) L’oreille interne
III. Neurophysiologie auditive
1) Transmission du son aux cellules ciliées
2) Voies auditives
3) Modulation
IV. Neurophysiologie du système vestibulaire
1) Intro
2) Les organes à otholites : accélération linéaire
3) Les ampoules : accélération angulaire
4) Transmission des messages nerveux
5) Intégration

Page 2 sur 40
L’objectif de ce cours est :
- de vous montrer que l’audition est un message sensoriel complexe (et que tout n’est pas encore
connu).
- Il faut connaitre le principe global de fonctionnement neurophysiologique de l’audition,
- et être sensibilisé au fait que le bruit et les problèmes auditifs constituent un problème de santé
publique majeur (ce qui sera en fait plus détaillé dans le cours de sémiologie de l’audition). Il y a de
plus en plus de cas de surdité précoce lié à nos habitudes de vie (écouteurs trop forts, boite de nuit,
concert). A partir d'une certaine intensité sonore on aura un trouble de l'audition liée à une
dégénérescence des cellules auditives, et malheureusement on n'est pas encore capable de les
régénérer.
I. Introduction
1) L’audition et la fonction de l’audition
L’audition correspond à la perception de l’énergie du son. Le son correspond à la partie audible du
spectre des vibrations acoustiques. Mais il ne faut surtout pas oublier que le spectre au complet des
vibrations acoustiques est beaucoup plus large que la petite partie de ce spectre que nous sommes capables
de percevoir.
Les ondes sonores sont perçues en tant que variations de la pression de l’air.
Ainsi tout ce qui est susceptible d’induire un mouvement de l’air sera audible. La réception se fait au
niveau des oreilles. Les nerfs auditifs vont atteindre le cerveau, transmettre l’info à l’aire dédiée et générer la
perception du son.
L’audition sert :
A communiquer essentiellement
A se reproduire: chez le cerf et chez la mésange la femelle choisit le mâle qui fait le plus de bruit
Alerter: le cri est différent pour alerter d’un danger chez les singes en fonction de l’animal qui les
menace (aigle, léopard, serpent).
A défendre son territoire: les phoques passent des heures à «chanter» pour empêcher les autres
males de s’approcher de leur territoire.
S’orienter et chasser: la chauve-souris, comme les baleines et les dauphins, grâce à
l’écholocalisation (fonctionnement de type sonar).

Page 3 sur 40
2) Les ondes sonores
Les ondes sonores correspondent à la progression de vibrations de l’air dues à l’alternance de zones de :
Haute pression (les molécules de l’air sont comprimées, concentrées)
Basse pression (les molécules de l’air sont raréfiées).
Le son correspond au final à une alternance de hautes et de basses pressions et tout cela va nous
donner une courbe sinusoïdale.
C’est la même chose quand vous faites vibrer un diapason : quand il ne vibre pas les molécules d’air qui
l’entourent sont réparties de façon homogène. Dès qu’il vibre : il va comprimer d’un côté les molécules d’air
et au contraire provoquer la raréfaction des molécules d’air de l’autre côté, et ce de façon séquentielle,
provoquant l’émission d’ondes sonores successives dans les différentes directions de l’espace (exactement
de la même manière qu’une pierre jetée dans un lac qui génère des clapots concentriques).

Page 4 sur 40
3) Les caractéristiques du son
D’un point de vue physique, l’onde sonore est représentée comme une courbe sinusoïde. On définit donc :
- L’amplitude
- La fréquence : qu’on définit comme étant l’inverse de la période
- Et l’intensité
Un son pur a une période nette, il a une courbe sinusoïdale parfaite.
Le son musical, s’il a effectivement une périodicité, n’a pas pour tracé une sinusoïde contrairement
au son pur puisqu’il correspond en fait à une succession spécifique d’harmoniques d’amplitudes
variables.
Le bruit n’a, quant à lui, ni périodicité, ni amplitude spécifique. Le bruit correspond à une
désorganisation totale.
Donc le son au final se définit par :
Sa fréquence (hertz) permet de définir les sons graves (fréquence basse) et des sons aigus (haute
fréquence).
Son intensité (décibel) ou puissance du son, est définie par l’amplitude des vibrations.
Son timbre (ou sa qualité)

Page 5 sur 40
Courbe audiométrique de l’oreille humaine : capacité à percevoir les différents sons (20Hz à 20kHz).
Ce qui est important de noter ici est que :
- l’oreille humaine n’a pas la même
capacité de percevoir toutes les fréquences
sonores à n’importe quelle intensité. A 40 db et à
20 Hz, par exemple, vous n’apercevrez de son. Il
faut atteindre une certaine intensité et une
certaine fréquence pour entendre quelque chose.
- La zone des conversations se situent entre
1 et 2 kHz.
- Notre spectre audible se situe entre 20Hz
et 20kHz.
La meilleure zone de perception se situe
entre 1 et 2 KHz – entre 1 à 120dB – car on a la
plus grande amplitude de détection des sons. Or
les voix sont entre cet intervalle qu’on nomme
zone conversationnelle. Si on dépasse les 150dB, on n’entend plus de son mais c’est un phénomène
nociceptif qui est déclenché.
On a donc défini des limites au champ auditif humain : la limite inférieure étant le seuil de détection du
son, et la limite supérieure de perception au-delà de laquelle l’intensité du son devient dommageable pour
l’oreille (les cellules ciliés étant détruites par des sons de trop forte intensité et n’étant pas remplacées, ces
sons provoquent une surdité).
Les sons inférieurs à 20 Hz ne sont pas perceptibles par l’oreille humaine et sont donc qualifiés
d’Infrasons. De même, les sons supérieurs à 20 000Hz ne sont pas non plus perceptibles et sont qualifiés
d’Ultrasons. Cette définition est, bien entendu, anthropocentrique puisque certains animaux sont capables de
les percevoir, voire d’en émettre.
Vous avez ici le spectre audible de l’Homme par rapport à celui d’autres espèces :
On voit ici qu'en fonction des animaux, ce spectre audible varie. Chez la baleine et le chien, la détection
sonore va de quelques Hz à quasiment 100.000 Hz alors que chez l'homme c'est de 20 à 20.000 Hz.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
1
/
40
100%