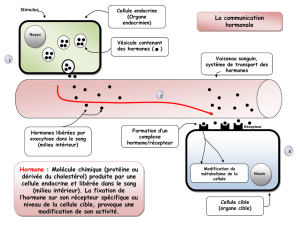
CONTROLES ET REGULATIONS Systèmes de communication et de

CONTROLES ET REGULATIONS
I. SYSTEMES DE COMMUNICATION ET DE REGULATION
A. NOTION DE CONSTANTE EN BIOLOGIE, DE MILLIEU INTERIEUR ET D’HOMEOSTASIE
Les constituants du sang ont des valeurs contantes : si on fait un prélèvement sanguin, on
observe que les concentrations de ces constituants sont égales dans le temps et d’un individu à
l’autre. Par ex : glycémie 1 g/l soit 0,5 mmol.l-1.
Chez le malade, certaines grandeur sont totalement différentes de la normale. On peut se
poser 3 questions en physiologie :
combien : environ 1 g/l
comment cette valeur est-elle atteinte
pourquoi : rôle pour la survie.
Cas du glucose dans le sang :
Combien ? Les valeurs varient dans certaines limites (au delà desquelles la santé n’est
plus maintenue).
Normalement : 1 g/l ou 5,5 mmol/l dans le sang et le LEC
Comment ? le volume du LEC est de 20 l :
masse de glucose = 20 g dans le milieu
intérieur.
Le pool de glucose du milieu intérieur sert
à alimenter les cellules musculaires et le méta-
bolisme.
Un exercice musculaire dépense 400 w (J).
Or, 1 g de glucose correspond à 17 kj. Le ren-
dement maximal est de 40 %. Donc 1 g de glu-
cose fournit 6,8 kj. D’où 400 w consomment
3,5 g de glucose chaque minute.
20 g de glucose sont épuisés totalement en 6 minutes.
Comme le taux sanguin reste le même, il y a renouvellement du glucose.
Le taux de renouvellement d’un constituant est la fraction du pool renouvelé, c’est à dire
entrant et sortant par unité de temps.
Le Pool étant la quantité de substance concernée dans le compartiment considéré.
Il est dans le cas présent de 3,5 g/min.
Il traduit le temps nécessaire pour qu’un volume (ou une masse) du constituant contenu
dans le compartiment y entre ou en sorte totalement.
C’est donc un rapport entre le contenu du compartiment et l’affluence ou la perte.
étude de ce qui permet la régulation de la glycémie.
Pourquoi ?
Pour permettre l’homéostasie c'est à dire la constance du milieu intérieur.
Claude Bernard : la survie d’un être vivant nécessite que ses cellules soient contenues
dans un milieu dont les constantes physico-chimiques soient stables (compatibles avec la sur-
vie) Ce maintien est permis par les organes qui alimentent ou qui éliminent les composants du
milieu intérieur.
(Homéostasie vient du grec homéo : stable et stase : position)
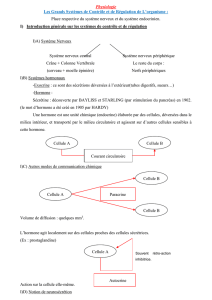
B. NOTION DE SYSTEME
Ex : système nerveux., cardio-circulatoire, respiratoire, ..
Un système est un ensemble arbitraire-
ment défini de tissus, d’organes, de cellules
qui exercent une fonction bien définie.
Un système (cardio-vasculaire) peut être
divisé en plusieurs systèmes étudiables.
.
20 g de glucose
D out
º
D in
º
20 g de glucose
sorties
SE
entrées système
physiologique
S = f(E)

3 grandeurs pour un système
On s’intéresse à ce qui sort du système (1ere étape : on sort de la boîte).
On analyse le fonctionnement du système (on entre dans la boîte) en le divisant en plu-
sieurs petits systèmes.
f = fonction de transfert ou loi du système (ce que cherche le physiologiste).
Par exemple : système rénal
sortie : urine (dépend de [urée] dans le sang et du débit du sang...)
entrée : sang
f : description du travail du système physiologique.
Les 3 grandeurs sont interdépendantes : la connaissance de 2 d’entre elles permet de connaître
la 3eme.
Le médecin observe les sorties anormales et fait des hypothèses de la cause du dysfonction-
nement diagnostic et prédiction de l’action thérapeutique par modification des entrées.
C. SYSTEMES THERMODYNAMIQUES FERMES ET EQUILIBRE
Notion d’entropie et d’énergie libre.
Un système fermé est un système isolé,
qui n’échange rien (ni matière, ni éner-
gie) avec son environnement.
L’énergie totale ET est la somme de
toutes les énergies contenues dans le sys-
tème.
Le système de départ contenait de l’énergie libre susceptible de se transformer en chaleur.
On appelle énergie libre d’un système la quantité d’énergie noble qu’il contient capable
de se transformer en une autre énergie (chaleur). L’énergie restante est égale à la variation
d’entropie x la température (Kelvin).
La caractéristique de la vie est d’organiser les systèmes pour leur faire baisser leur entro-
pie et leur faire gagner de l’énergie libre qui peut se faire par une hausse de l’entropie dans le
milieu extérieur gain d’organisation dans le système physiologique. On parle de négentro-
pie. D. SYSTEMES THERMODYNAMIQUES OUVERTS
Il existe deux mécanismes fondamentaux qui peuvent expliquer la constance.
1. Etat stable
L’état stable, c’est la constance d’une valeur atteinte par la mise en jeux de certaines
lis physico-chimiques
Exemple du bassin :
h tend spontanément vers une certaine
valeur stable.
Si il n’y a pas d’eau dans le bassin,
Dout = 0. Or, Din est constant. Plus l’eau
s’accumule, plus Dout augmente.
Quand h augmente, Dout tend à équili-
brer Din h tend vers un état stable.
Beaucoup de grandeurs (cf glucose)
sont comparables à ce bassin.
.
loi d’action de masse : plus la teneur augmente plus le débit de sortie augmente
énergie
totale énergie
libre variation
d'entropie température
(Kelvin)
ET = EL + s . T
origine : système ordonné
(non homogène) le système gagne en homogénéité
(désordonné : gain en entropie)
S = Q/T
quantité
de chaleur en K
D in
ºentrée
D out
º
h
sortie

On appelle ainsi des états stables qui n’impliquent aucun phénomène particulier si ce
n’est des lois physico-chimiques.
Si on fait baisser le niveau (avec un seau d’eau), spontanément, Dout diminue jusqu’au
retour spontané à l’équilibre.
Ceci est amélioré par le développement de systèmes spécifiques qui augmentent la ca-
pacité de maintenir le niveau constant.
2. Régulation
Quand le niveau baisse, l’entrée aug-
mente : c’est une rétro-action ou feed-back.
Quand la rétro-action tend à corriger la
cause qui l’a fait naître, c’est une rétro-action
négative : son effet s’oppose à la cause qui l’a
fait naître.
Une ante-action positive corrigeant la
cause qui l’a fait naître peut agir au niveau de
: la sortie : feedforward positif.
l’entrée : feed back négatif
On peut imaginer que quand le niveau baisse, l’admission se ferme et le bassin va se
vider encore plus : un tel phénomène vidant complètement le bassin est un feed-back positif.
L’action sur les entrées a pour effet d’augmenter l’amplitude de la réponse.
Un feed-back positif = feed forward négatif.
Il en existe en physiologie : dépolarisation membranaire, décharge ovulante.
Cependant, le feed-back négatif est la régulation la plus fréquente.
Les avantages du feed-back :
améliore la performance des états stables
augmente la capacité à maintenir les grandeurs constantes.
a) Grandeur régulée
C’est la grandeur maintenue constante. Si l’on observe une grandeur constante, la
grandeur régulée peut être un de ses corrélats.
La grandeur régulée est la grandeur qui est mesurée par des capteurs;
Une grandeur physiologique régulée est la température (il existe des cellules qui cap-
tent la température).
Souvent, on ne sait pas comment sont mesurées certaines valeurs, donc on ne sait pas
si il y a régulation.
b) Capteurs
Une régulation ne peut se faire sans mesures biologique, et ces mesures sont faites par
des capteurs multiples qui renseignent le cerveau.
Ici, le capteur est le flotteur.
Le capteur doit envoyer l’information au moyen d’une boucle informationnelle :
boucle de rétro-action, ou boucle de feed-back. Dans l’organisme, il existe beaucoup
de boucles (chimiques ou neuronales).
c) Valeur de consigne : set point
C’est une information intrinsèque au système qui détermine la valeur régulée On
l’appelle valeur de consigne. Si on raccourcit b, on va changer l’équilibre de h en h’>h. La
nature biologique des valeurs de consignes est très variable : seuil d’activation d’un récepteur,
perméabilité membranaire...
En général, une modification de la valeur de consigne est responsable d’une patholo-
gie. d) Comparateur
Seule, la valeur de consigne ne sert à rien, elle doit être comparée à la valeur mesurée,
pour voir s’il y a une ou plusieurs différence pour qu’il y ait ou non une action réparatrice.
Si un écart existe, le système va déclencher un processus de régulation.
D in
ºentrée
D out
º
h
= f(h)
axe
flotteur
tige métallique
admission
b

e) Signal d’erreur
Il n’y a pas de régulation sans signal d’erreur : pour qu’il y ait une correction, il faut
qu’il y ait un écart.
Régulation veut dire oscillations autour de la valeur de consigne.
Si on dépasse la valeur de consigne, le signal d’erreur déclenche une réaction. Toute
régulation suppose l’existence d’un signal d’erreur.
f) Gain d’une régulation
Il correspond au rapport entre l’écart observé sur la sortie du système en réponse à une
perturbation donnée, et en l’absence de toute régulation, sur le signal d’erreur avec la même
perturbation, lorsque la régulation fonctionne correctement.
Plus une régulation a un gain élevé, moins la marge de variation est grande.
g) Représentation schématique d’une régulation
h) Loi du système
C’est la relation qui existe entre le signal d’erreur et la réponse correctrice.
Régulation proportionnelle
Si la perturbation ne varie pas, il y a toujours un signal d’erreur dans le cas d’une ré-
ponse proportionnelle.
réponse proportionnelle = réponse phasique.
Réponse dérivée : réponse dont l’amplitude est proportionnelle à la dérivée par rapport
au temps du signal d’erreur.
la réponse corrective n’existe que si la vitesse de variation existe.
correction uniquement quand la variable régulée s’éloigne de la valeur de consigne.
réponse de type tonique accélère l’efficacité des réponses. Cela accroît la vitesse de
correction. Si on a eu un retour vers la valeur normale, la réponse est négative : rôle de frein
de la réponse dérivée.
Souvent, les deux réponses se cumulent : réponse proportionnelle et dérivée : réponse
tonico-phasique.
3. Contrôle : différence entre variable régulée et contrôlée.
La pression sanguine est une variable régulée.
+-
valeur de
consigne signal
d'erreur LOI
capteur valeur de sortie
boucle
informationnelle
comparateur
nerf
centre
bulbaire
tenso-récepterur
aortique
temps
valeur de consigne
transitoire
signal d'erreur
perturbation
réponse
régulatrice
proportionnelle
dérivée
proportionnelle
+ dérivée
continue
réponse dont l'amplitude est
proportionnelle au signal d'erreur

L’élément régulateur est l’accélération cardiaque. La modification de la fréquence
cardiaque est la variable contrôlée : c’est la variable au moyen de laquelle l’organisme assure
la régulation.
4. Servomécanisme et mécanisme d’homéorheusie
L’homéorheusie est l’adaptation dans le temps de l’homéostasie aux besoins de
l’organisme.
La température du corps est régulée à 37°c. Mais la température s’élève chez la femme
après l’ovulation : la valeur de consigne n’est pas tout à fait la même en phase pré et post ovu-
latoire. De même, chez tout individu, la température est un peu plus élevée le soir que le ma-
tin. Exemple du pilote automatique
Diabète : insuffisance d’insuline (elle permet de faire entrer le glucose dans la cellule).
La concentration du glucose sanguin augmente pour augmenter le gradient de concen-
tration.
Différents types de régulations biologiques :
régulation autonome : se fait de manière réflexe et simultanée à d’autres régula-
tions. régulation comportementale : nécessité d’un ordre de priorité : on a soif avant
d’avoir faim.
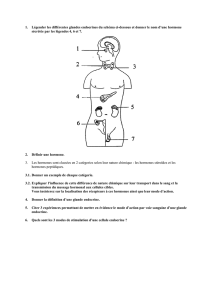
II. LES GRANDS SYSTEMES DE CONTROLE ET DE REGULATION DE L’ORGANISME :
PLACE RESPECTIVE DU SYSTEME NERVEUX ET DU SYSTEME ENDOCRINIEN
A. INTRODUCTION GENERALE SUR LES SYSTEMES DE CONTROLE ET DE REGULATION
La physiologie est la science des régulations.
2 éléments sont importants :
information : récepteurs,...
commande
Il faut nécessairement une circulation de l’information par l’un des deux modes :
système nerveux
système hormonal
système nerveux fréquence cardiaque
système contrôlé système régulé
commande
pression artérielle
réglage
commande
mécanisme d'homéorheusie
consigne +-
ENTREE
controleur action de
contrôle système
passif
capteur
signal de
rétroaction
(sb)
consigne
- sb hsortie
énergie perturbation
système contrôleur système contrôlé
 6
7
8
9
10
11
12
13
14
15
6
7
8
9
10
11
12
13
14
15
1
/
15
100%