Ronéo

1
Physio de la fonction visuelle
I. Rappel d'anatomie:
L'espace objet est séparé en 2 parties temporales (extérieures) et 2 nasales (pour chaque oeil)
formant le champs visuel
Deux yeux fonctionnels (c'est pas si fréquent) nous donnent une vision binoculaire permettant de
distinguer les reliefs important pour l'appréciation des distances.
Voies de conduction visuelles :
• Deux nerfs optiques (schéma: 1) constitués des axones des cellules
ganglionnaires de la rétine nasale et temporale
• Chiasma: (2) en contact avec l'hypophyse (pouvant appuyer sur le
chiasma si tumeur), décussation partielle: croisement des axones
des provenant de la rétine nasale, les temporales étant directes: ça
permet un regroupement des informations par hémichamp (en gros
ce que chaque oeil voit à droite part à droite et inversement)
• Tractus optique, rétro chiasmatique: réunion des nerfs (4)
• Relais synaptiques aux Corps Géniculés latéraux: 2 zones
synaptiques (1 droit et 1 gauche) permettant une amplification
notamment des structures maculaires (ce sera plus clair après avoir compris la suite)
• Radiations optiques (6) se terminant de part et d'autre de la
scissure calcarine au niveau du cortex visuel primaire
• Cortex visuel primaire et Nombreuses aires visuelles associées
Stimulus: Stimulation qui aboutit à une sensation.
En optique, le stimulus est la lumière et la sensation est la perception visuelle.
La lumière du jour peut être décomposé par un prisme en un spectre continu où chaque longueur
d'onde correspond à une couleur. Or, le spectre du visible pour l'homme s'étend sur une gamme
restreinte de longueur d'onde: de 400 à 700 nm. Pourtant on ne détecte que 150 tonalités (de 2 en
2nm) de lumière spectrale. Si on voit plus de couleurs c'est parce qu'on a des nuances (clair ou
foncé) ou des différences d'intensité...
Cependant, l'oeil est capable de mettre en activité le système visuel selon une gamme étendue de
niveau lumineux. En effet on perçoit à la fois, de faibles niveaux lumineux (dits niveaux lumineux
scotopiques), on parle de vision de nuit (phare de voitures, éclairage faible): perception de gris clair
à très foncé, mauvaise acuité visuelle
Fort niveau lumineux (dits niveaux lumineux photopiques), on parle de vision de jour: perception
en couleur, bonne acuité visuelle (nous sommes donc des animaux diurnes à la différence des
rongeurs...)

2
II. La Rétine
La rétine est le récepteur des images de l'espace objet. Il y a transmission de la stimulation à travers
les structures transparentes (cornée, humeur aqueuse, pupille (diaphragme qui régule la quantité de
lumière entrante), cristallin, humeur vitrée, et enfin la rétine). Le but c'est que, pour voir clair, le
plan focal soit confondu avec le plan de la rétine (cf cours de P1 pour les plus motivés)
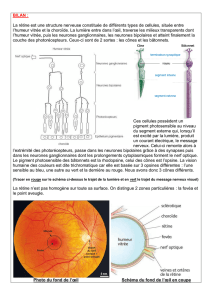
A) fond d'oeil
La rétine est visible si le fond de l'oeil est éclairé à travers la pupille. On voit une structure rosée
avec une zone plus foncée au centre: la fovéola (point d'impact de l'axe visuel: l'image d'un objet
qu'on regarde se projette exactement sur elle). La papille est une zone plus claire qui forme le lieu
de passage des axones des cellules ganglionnaires formant le nerf optique. Mais il n'y a pas de
rétine à cet endroit et du coup pas de vision. Les artères sont vues plus claires que les veines.
Autour de la fovéola, il y a une zone vue sous un angle de 10-15° qui correspond à la rétine centrale
qui est liée au champs visuel central (zone où on voit au top de jour, si elle est normale). Autour:
rétine périphérique (ou pôle postérieur): champ visuel périphérique.
B) Architecture fonctionnelle
- Niveaux lumineux photopiques: la rétine périphérique est une zone d'alerte. Elle déclenche le
mouvement des yeux ou de la tête pour amener l'image en zone centrale (ex: Si quelqu'un fait le
fou-fou à coté, on le verra mais pas bien alors on va tourner la tête pour le mettre en rétine centrale)
- Niveaux lumineux scotopiques: la rétine centrale n'a pas ou peu de vision. La rétine périphérique a
une mauvaise acuité visuelle mais c'est la seule à percevoir de faibles niveaux lumineux.
On trouve deux systèmes (à ne pas confondre avec au dessus):
- le système photopique (= système des cônes), est lié aux niveaux lumineux photopiques:
responsable de la vision de jour, en couleur
- le système scotopique (= système des bâtonnets) est lié aux niveaux lumineux scotopiques:
responsable de la vision de nuit
La rétine est l'organe de réception des images et de transduction (transformation des signaux
lumineux en signaux électro-physiologiques interprétables par le cerveau)
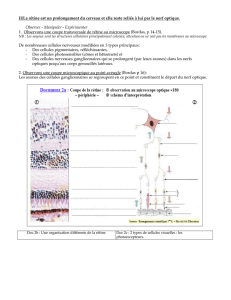
C) Organisation
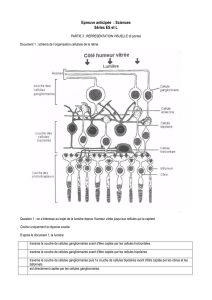
- Rétine périphérique:
3 étages fonctionnels qui reposent sur l'épithélium
pigmentaire (au dessus du schéma):
- 1: Etage des Photorécepteurs : cônes et bâtonnets (le +
distale: postérieur par rapport à l'entrée de la lumière)
/ Couche Plexiforme Externe: cellules Horizontales
(tangentielles), zone de synapse entre l'étage 1 et 2/
- 2: cellules bipolaires de cônes et de bâtonnets (différentes)
/ Couche plexiforme interne (sous couches a et b):
cellules amacrines (tangentielles, rôle trophique)/
- 3: cellules ganglionnaires dont les axones forment le
nerf optique: amyéliniques dans la rétine, myélinisation à
partir de la lame criblée au niveau de la papille

3
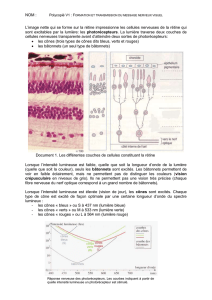
- Rétine centrale:
Portion de quelque degrés autour de fovéola. C'est une exception rétinienne car il n'y a qu'un seul
type de cellules (cônes) au niveau de la fovéola: la zone de 350µm, vue sous un angle de 2° et
avasculaire. Ainsi, il n'y a pas à traverser les 3 étages: les cônes sont atteints directement par la
lumière. Seul l'homme à une fovéola
On en a de belles images par la technique OCT (Optical Coherence Tomography) qui donne de très
bonnes résolutions sur quelques millimètres de profondeurs et est donc tout à fait adapté à l'optique.
III. Les photorécepteurs
A) Caractéristiques des photorécepteurs (cônes et bâtonnets):
- Ils comprennent une zone synaptique, un article interne et un article externe: partie supérieure en
forme de cône ou de bâtonnet, contenant les photopigments fixés dans la membrane. Ils
interceptent les photons et permettent la transduction. Les bâtonnets contiennent de la Rhodopsine,
les cônes contiennent le pigment S, M ou L mais jamais de mélange. On a donc 3 cônes différents:
les cônes L, les cônes M et les cônes S
- Nombre: 100 millions de bâtonnet; 4,5 millions de cônes en fonction des gens, mais on a toujours
90% de photopigments L et M et 10% de S
- Les cônes sont régulièrement répartis entre les bâtonnets sur l'ensemble de la rétine sauf à la
fovéola où il n'y a pas de bâtonnets et donc uniquement des cônes (pas de vision précise la nuit)
- Densité: Les cônes sont beaucoup plus denses à la fovéa qu'à la périphérie. Mais la densité de
cellules est supérieure à la périphérie en partie parce que les bâtonnets sont beaucoup plus petits et
donc il y en a plus par unité de surface.
B) Propriété
Transduction: Transformation d'un signal lumineux en signal électro-physiologique.
L'étape initiale est l'absorption des photons et le résultat est la variation de l'état de polarisation des
photorécepteurs transmise aux cellules bipolaires puis aux cellules ganglionnaires qui transforment
variation de polarisation en Potentiels d'action pour transmettre l'information.
Le mécanisme est le même pour les cônes et les bâtonnets.
- Dans l'obscurité : les photorécepteurs sont dépolarisés. Il y a donc une libération importante de
neurotransmetteurs (dans ce cas le Glutamate).
- A la lumière: les photorécepteurs sont hyperpolarisés. Il y a donc moins de libération de Glutamate
au niveau de la synapse avec la cellule bipolaire. En fait, avec l'arrivée du photon, il y a une cascade
de boucles d'activation qui fonctionne avec une protéine G. On aboutit à la fermeture de canaux Na+
qui cause l'hyperpolarisation.
Graphe 1 Graphe 2

4
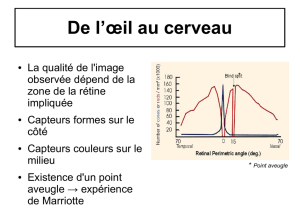
Pour des niveaux lumineux scotopiques (Graphe 1), seule la Rhodopsine est capable d'absorber les
photons donc seuls les bâtonnets fonctionnent. Il y a donc codage des longueurs d'onde en fonction
des différentes probabilités d’absorption des photon par la rhodopsine. De plus, on voit qu'avec
de faibles niveaux lumineux, les longueurs d'ondes supérieures à 650nm ne seront pas vu.
On voit d'autant plus clair que la probabilité d'absorption est élevé, maximale à 510nm. Les deux
longueurs d'onde indiqués seront vu avec la même intensité...
Pour des niveaux lumineux photopiques (Graphe 2), seuls les cônes codent. Le photopigment S a
une probabilité d'absorption maximale à 420nm, le M à 530nm et le L à 600nm. (ds l'ordre: ça fait small...^^)
On remarque qu'avant 400nm et au-delà de 700nm, on n'a pas de photopigments.
Le problème c'est qu'en 445 et 475nm, les pigments S ont la même probabilité d'absorption, ils
perçoivent donc ces deux longueur d'onde comme identique (même polarisation). Mais la
probabilité d'absorption en M et L sont différentes pour ces longueurs d'onde. C'est ce qui fait qu'on
a une sensation différente, c'est à dire que l'on voit des couleurs différentes.
En fonction de la culture, la gamme des couleurs est différente. Par exemple, les Inuits ont plus
d'une centaine de noms de blancs différents. Pareil pour le marron en Afrique.
C) Pathologies
- Dyschromatopsie: déficience de la vision des couleurs généralement due à l'incapacité de
distinguer deux longueurs d'onde qui ont la même probabilité d'absorption (absence de cônes M...)
- Daltonisme: absence de fonctionnement de cônes M, L ou des deux (mais il en reste au moins 1)
- Achromates: privés de l'utilisation des cônes; ne perçoivent pas les couleurs
IV. Voies des cônes et des bâtonnets
A) Voie des cônes (K, P et M)
On va voir se qui se passe après le photorécepteur, c'est à dire aux 2e et 3e étage rétinien. On
rappelle que les cônes sont suivis des cellules bipolaires puis des cellules ganglionnaires.
Il faut distinguer la voie des cônes M et L qui est complexe: ce sont les voies P (comme Parvo) et
M (comme Magno), nommées ainsi car elles finissent aux couches parvocellulaire et
magnocellulaire des corps géniculés latéraux et la voie des cônes S, beaucoup plus simple: c'est la
voie K (comme Konio) qui se termine à la couche inter laminaire des corps géniculés latéraux.
Ces voies ne fonctionnent qu'avec des Niveaux lumineux photopiques
- La voie P concerne les cônes L ou M. Chacun est lié à 2 cellules bipolaires de type naines On et
Off, elles même reliées aux cellules ganglionnaires naines On et Off
Les cellules bipolaires naines On vont se dépolariser et les Off s'hyperpolariser en cas de
stimulation.
- La voie M concerne aussi les cône L et M. Chacun est lié à 2 cellules bipolaires de type diffuses
On et Off, elles même reliées aux cellules ganglionnaires parasols On et Off. La différence est que
les cellules bipolaires entre en contact indifféremment avec les cônes L ou M alors que les
bipolaires naines séparent bien les cônes L des M. De plus, plusieurs cellules bipolaires sont
connectés à la même cellule ganglionnaire.
Au centre de la rétine, la voie P lie un cône à 2 cellules bipolaires. Il y a une divergence et donc une
amplification de l'information, ce qui pourrait expliquer la bonne acuité visuelle de cette zone. Pour
la voie M, plusieurs cônes sont liés à une seule bipolaire, il y a donc convergence de l'information.
En périphérie, les voie P et M sont toutes les deux convergentes. Il y a donc un certain degré de
compression de l'information. Il y a donc une moins bonne performance: pas d'amplification.
On comprend ainsi pourquoi les gogols qui ont regardé le soleil pendant l'éclipse et qui ont grillé

5
leur fovéola ne voyaient plus rien pour seulement quelques degrés de brûlés.
- La voie K concerne les cônes S. Chacun est lié à 1 cellule bipolaire de cône S de type On, elle
même reliée à 1 cellule ganglionnaire bistratifiée qui reçoit les informations de S, mais aussi de L et
de M. En fait 5 cônes sont liés à 1 cellule bipolaire, il y a donc convergence. Les cônes S ont un rôle
fondamental pour la vision des couleurs mais sont peu nombreux et fragiles (ce sont donc les
premiers touchés lors de pathologies).
Voie P Voie M Voie K
Les voies P (80%), M (19%) et K (1%) forment 100% du nerf optique: que reste-t-il pour les bâtonnets?
B) Voie des bâtonnets
C'est une voie parasite. Rôle visuel mineur (vision de nuit). Cependant ils ont un rôle majeur pour le
trophisme des cônes. Ainsi, s'il y a un dysfonctionnement des bâtonnets (héméralopie: absence de
vision de nuit), on aura un retentissement sur le fonctionnement des cônes. On a donc de plus en
plus de thérapies visant la survie des bâtonnets pour toucher les cônes.
La voie des bâtonnets est une voie parasite car, arrivant les derniers dans l'embryogénèse, ils se
connectent directement sur la voie P déjà en place. Ils l'utilisent quand les cônes ne fonctionnent pas
(milieux scotopiques). 35 à 40 bâtonnets se connectent sur 1 cellule bipolaire des bâtonnet qui
transmettent l'information aux cellules amacrines AII qui se branchent sur les cellules
ganglionnaires naines On et Off. S'ils n'utilisent pas la voie P, ils ont leur rôle trophique.
Conclusion
Toute altération du champ visuel central entraîne des modifications importantes de l'acuité visuelle
et de la vision des couleurs. C'est d'ailleurs souvent le premier signe d'altération de la rétine centrale.
Toute altération du champ visuel périphérique entraîne: de jour des modifications de la perception
des mouvements, et de nuit des modifications de la perception des faibles niveaux lumineux
Neuropathie optique: Altération du nerf optique, touche essentiellement la voie P. Elle peut être
d'origine toxique (alcool où quand on veut frimer avec un terme médical: exogénose),
médicamenteuse (éthambutol: antituberculeux), compressive (glaucome) ou dégénérative
(démyélinisation)
1
/
5
100%