Physiologie musculaire

1
Physiologie musculaire
I) Généralités
- Les muscles représentent 40 % du poids du corps et comprennent les muscles striés
squelettiques et cardiaque et les muscles lisses.
- Le muscle convertit une énergie chimique en énergie mécanique avec un rendement élevé de
50 à 60 %.
- Ils sont organisés selon des systèmes de levier qui permettent une amplification de la force
développée et du déplacement.
- Le muscle est formé de l'assemblage de faisceaux eux-mêmes rassemblant les fibres
musculaires. Chaque structure est entourée d'un tissu conjonctif.
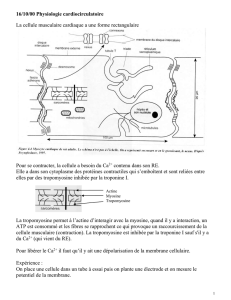
II) Structure de la cellule musculaire striée
- La cellule musculaire est une cellule géante multinuclée, remplie de myofibrilles; c'est la
cellule qui constitue la fibre musculaire.
- Les myofibrilles comprennent trois types de protéines :
- Les protéines contractiles : actine et myosine.
- Les protéines de soutien : -actinine, titine, vinculine, myomésine.
- Les protéines contrôlant la contraction : tropomyosine et troponines C, I et T.
- Vu en microscopie optique, la fibre musculaire présente un aspect strié, qui lui donne son
nom et est dû à l'organisation des myofibrilles.
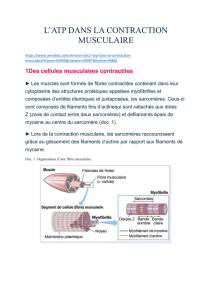
Le filament épais :
- Il est constitué de l'assemblage de myosine, protéine de 510 kDa, constituée de :
- Deux chaînes lourdes enroulées en double hélice et terminées par deux renflements
S1 et comprenant le site de liaison avec l'actine et le site de fixation et d'hydrolyse de l'ATP.
- Quatre chaînes légères.
Le filament fin :
- Sa composition est plus complexe ; il est formé de l'assemblage d'actine, de tropomyosine et
de troponines.
- L'actine F est une double hélice d'actine G.
- La tropomyosine recouvre au repos les sites de fixation de la myosine.
- Les troponines sont de trois sortes : la troponine T (30 kDa) liée à la tropomyosine, la
troponine I (30 kDa) qui empêche l'interaction myosine - actine et la troponine C (18 kDa) qui
contient les sites de liaison pour le Ca2+
Cet ensemble de filaments est maintenu par des protéines de soutien et leur agencement se fait
sous forme de l'unité contractile, le sarcomère.
Le sarcomère est un segment de myofibrilles compris entre deux stries Z.
- Zone H : filament épais sans tête de myosine
- Bande A : filament épais sur toute sa longueur
- Bande I : filament fin
- Ligne M : myomésine
- Strie Z : -actinine, limite du sarcomère
- Lien entre -actinine et membrane plasmique : vinculine et dystrophine
- Support de la myosine : titine

2
Des éléments très divers participent à la contraction musculaire :
- Les myofibrilles contiennent les structures contractiles et élastiques.
- Le sarcoplasme et les mitochondries sont les structures métaboliques nécessaires au
fonctionnement.
- Le sarcoplasme et les tubules transverses servent à la transmission nerveuse du message
activateur.
- Les gaines conjonctives représentent les structures qui transmettent les forces.
III) Aspect microscopique de la contraction musculaire
1) Cycle des interactions
- Le principe de la contraction musculaire repose sur le glissement des filaments l'un sur
l'autre et le moteur qui permet ce glissement est lié à l'interaction entre les têtes de myosine et
l'actine par une liaison faible non covalente ATP dépendante.
- Le mécanisme générant le mouvement n'a pas encore été totalement élucidé et plusieurs
théories ont été élaborées successivement. La théorie du cliquet brownien pourrait expliquer
la variabilité du saut de la tête de myosine sur le filament d'actine.
- Cycle des interactions :
1. Les têtes de myosine contiennent ADP + Pi : pas d'interaction car la tropomyosine masque
les sites.
2. En présence de Ca2+, il y a changement de conformation de la troponine I et déplacement
de la tropomyosine.
3. Le déplacement de la tropomyosine entraîne la formation de l'actomyosine : évacuation de
ADP + Pi, changement de conformation et glissement des fibres d'actine.
4. En présence d'ATP, une molécule d'ATP vient se loger dans la poche et est hydrolysée :
retour de la tête à sa position initiale.
5. La position de la tropomyosine détermine l'étape suivante
- Si la tropomyosine cache les sites de fixation à l'actine : pas d'interaction
- Si les sites sont découverts : interaction
- L'hydrolyse de l'ATP permet le retour de la tête de myosine à sa position de départ
- Sans ATP, le muscle reste contracté
- L'hydrolyse de l'ATP se fait par une activité ATPasique liée à la myosine

3
2) Régulation
- La régulation de ces cycles se fait par l'intermédiaire des flux calciques.
- A faible concentration de Ca2+ (10-7 M / L), troponines et tropomyosine s'interposent entre
l'actine et la tête S1.
- A forte concentration de Ca2+ (10-5 M / L), la troponine C fixe le Ca2+, ce qui entraîne une
modification de la configuration spatiale de la troponine I et de la tropomyosine, rendant
l'interaction possible.
- Le nombre d'interactions actine – myosine dépend du nombre de troponine C ayant fixé du
Ca2+.
- Le taux d'ATP est maintenu élevé et constant grâce au système de la créatine phosphate :
Créatine P ATP
créatine transférase
3) Couplage électromécanique
- Trois paramètres mesurables : - Le potentiel de membrane
- La concentration de Ca2+
- La force générée
- La membrane subit une
dépolarisation puis une repolarisation
progressive.
- La [Ca2+] augmente jusqu'à un pic
puis diminue.
- La force générée présente également
un temps de latence et augmente
progressivement même après la fin du
signal et du pic calcique.
4) Rôle de la durée de dépolarisation
Il existe une relation de proportionnalité entre force et [Ca2+]
- DF / dt = k1 . [Ca2+]i
- F(t) = k2 0t [Ca2+]i . t . dt

4
5) Rôle de l'intensité de dépolarisation
- On utilise un système de potentiel imposé qui permet d'étudier les effets des variations
d'intensité de stimulation.
- Une dépolarisation entre –50 mV et –
20 mV déclenche une contraction.
- La force de contraction est
proportionnelle à la dépolarisation
jusqu'à – 20 mV.
- A partir de – 20 mV la force
développée devient constante.
- Le muscle ne répond pas à une loi du tout ou rien
- La modulation de la réponse mécanique se fait en fonction :
- de la durée de la dépolarisation
- de l'intensité de la dépolarisation
IV) La plaque motrice et la triade
1) La plaque motrice
- La jonction neuro musculaire se fait par l'intermédiaire de récepteurs canaux à l'ACh dans le
sarcolemme.
- Le potentiel de plaque motrice déclenché par la fixation du neurotransmetteur est un
potentiel électrotonique ; il ne présente pas les mêmes caractéristiques que le potentiel
d'action axonal.
- Il ne s'autorégénère pas, mais diminue avec la distance.
- Il dépend des propriétés de conduction de la membrane.
- Chronologie de l'excitation musculaire :
- Arrivée du PA neuronal.
- Ouverture des canaux Ca2+ de type L.
- Libération d'ACh.
- Fixation de l'ACh sur le récepteur canal.
- Entrée de Na+ et sortie de K+ : modification de la conductance de la membrane.
- Dépolarisation de la membrane.
- La dépolarisation due à l'ouverture du canal doit atteindre un certain seuil pour permettre le
déclenchement du PA musculaire (- 48 mV).
- Le couplage entre la dépolarisation de membrane et l'ensemble des myofibrilles se fait par
l'intermédiaire d'une structure cellulaire, la triade.

5
2) Structure et fonctionnement de la triade
- Elle est formée par l'association d'un tubule transverse T avec des récepteurs DHP et de deux
citernes du réticulum sarcoplasmique comprenant les récepteurs à la ryanodine en surface et la
caléquestrine à l'intérieur.
- Le fonctionnement de la triade peut être mis en évidence par deux expériences :
- Dépolarisation localisée au tubule T
L'application d'un potentiel entraîne
un raccourcissement du sarcomère
sous jacent à la triade.
- Voltage imposé avec concentration variable en Ca2+ et Na+
- Le couplage est très localisé au niveau de la triade
- Il ne nécessite pas de Ca2+ extracellulaire.
- Il nécessite une concentration normale de sodium extracellulaire.
- Il utilise les stocks de calcium du réticulum.
- Il est lié à la présence de deux types de récepteurs :
- Récepteurs aux DHP voltage dépendant
- Récepteurs à la ryanodine
- Les canaux calciques du réticulum sarcoplasmique sont à la fois autoentretenus par le Ca2+
et inhibés par le Ca2+.
- La durée du signal calcique très brève et proportionnelle à la durée et à l'intensité de la
dépolarisation.
 6
6
1
/
6
100%