Embryologie du 4 mars

1
Embryologie du 4 mars.
Rappel
A la fin de S3, l’embryon mesure 1mm. Sa vésicule vitelline en dessous, sa vésicule
amniotique au dessus. L’ensemble flotte dans le cœlome externe et il y a le chorion composé
de la somatopleure et du trophoblaste et tout autour c’est la paroi utérine / l’endomètre utérin.
L’ensemble de l’embryon et de ses annexes est totalement englobé dans la muqueuse utérine.
La quatrième semaine.
A ce stade, on est presque plan. On est pas délimité par rapport aux vésicule vitelline et
amniotique alors qu’à J28 l’embryon est complètement délimité et il est raccordé simplement
au chorion par le cordon ombilical qui s’est délimité.
J21
C’est la poursuite de la neurulation. On en était resté au stade de la gouttière neurale. La
fermeture du livre va se poursuivre jusqu’à ce que les berges de la gouttière neurale se
rapprochent et fusionnent. Cette fusion se fait initialement au niveau environ du 4ème somite.
Cette fusion a plusieurs conséquences :
- au niveau des crêtes neurales qui était ce contingent de cellules à la jonction entre la
gouttière neurale et l’ectoblaste, elles se dissocient du tube neural et migrent activement au
sein du mésenchyme embryonnaire tout en proliférant. Au bout du compte ce petit contingent
de cellules a une telle activité de prolifération qu’il représente environ 10% des cellules de
l’organisme.
- la fermeture du tube neural est responsable de la fermeture de l’ectoderme (ectoblaste =
épiblaste) qui recouvre maintenant la face dorsal de l’embryon et les cellules des crêtes
neurales se retrouvent entre ectoblaste et tube neural, situation à partir de laquelle elles vont
migrer tout en proliférant.
Cette fusion va se poursuivre à la fois en direction craniale et caudale. A ce stade, la lumière
du tube neural communique avec la cavité amniotique au niveau des neuropores antérieurs et
des neuropores postérieurs. Ces neuropores sont des endroits où la gouttière neurale n’est pas
encore complètement fermée. La fermeture du neuropore antérieur se fait entre 24 et 26 jour.
La fermeture du neuropore postérieur entre 26 et 28 jour. Lorsque le neuropore antérieur ne se
ferme pas c’est une anencéphalie (pas de développement d’encéphale). Lorsqu’il y a non
fermeture du neuropore postérieur c’est responsable du spina bifida (grave à moins grave).
Dès le début de S4, le tube neural comprends une portion craniale large à l’origine de
l’encéphale et une portion caudale étroite à l’origine de la moelle épinière.
J23
L’extrémité céphalique se fléchit en direction ventrale et c’est la survenu de cette inflexion
qu’on appel la courbure céphalique qui permet de distinguer les 3 grandes régions de
l’encéphale :
- Au milieu de la courbure céphalique c’est l’encéphale moyen ou mésencéphale.
- En avant l’encéphale antérieur ou pro encéphale.
- En arrière l’encéphale postérieur ou rhombencéphale qui est suivit de la futur moelle
épinière.
Au niveau de ces segments encéphaliques on distingue des renflements qui sont des
subdivisions de ces différents segments. Ces subdivisions sont les neuromères. Il y en a 2 au
niveau du pro encéphale, 2 au niveau du mésencéphale, 9 au niveau du rhombencéphale. Un
de ces neuromères rhombencéphalique est intermédiaire entre le rhombencéphale et le

2
mésencéphale et les 8 neuromères suivant sont appelés les rhombomères. Au niveau crânien,
la segmentation du tube neural et des dérivés du mésoblaste para axial (somitomères et
somites) ne se fait pas en parallèle. En effet, les 7 premiers rhombomères sont au même
niveau que les somitomères crânien 4 à 7. Le 8ème rhombomère est au même niveau que les 4
somites occipitaux. A l’inverse, au niveau de la moelle la segmentation va se faire
parallèlement c’est à dire qu’il va y avoir émergence d’un nerf spinal au niveau de chaque
somite. Au niveau de la moelle l’association des dérivés d’une paire de somites et des dérivés
du tube neural de même niveau est appelé un métamère. Le tube neural résultant de la
fermeture du neuropore postérieur se termine à hauteur de la 31ème paire de somites : 2ème
sacrée, c’est à ce niveau là qu’émergera la 27ème paire des nerfs spinaux.
Il y a un tube neural qui se forme en regard des somites les plus caudal (1997/98) : la
formation la plus caudal du tube neural provient d’un processus de neurulation secondaire par
différenciation neural à partir de la partie médiane du mésoblaste du bourgeon caudal en
arrière du nœud de Hensen. Ce tube neural secondaire fusionne avec le tube neural primaire.
Donc maintenant on a un tube neural complet.
- Devenir des somites (structures éphémères) : A peine sont elles individualisées qu’elles se
subdivisent pour donner les compartiments somitiques. On en distingue 3 qui sont nommés
d’après leur position au début de S4 sur un embryon encore plan. On distingue le cadran
médio ventral, le cadran médio dorsal et la moitié latérale. Ultérieurement, du fait de la
plicature de l’embryon la position dans l’espace de ces différents compartiments va se trouver
modifié mais néanmoins on continue toujours à les identifier d’après leur position initiale.
- cadran médio ventral : forme le sclérotome dont les cellules subissent une
transformation épithélio mésenchymateuse et migrent en direction du tube neural et de la
chorde. Les cellules les plus ventrales (qui migrent autour de la chorde) contribuent à la
formation du corps vertébral. Les cellules les plus dorsales qui migrent autour du tube neural
contribuent à la formation de l’arc vertébral. Les sclérotomes sont des structures segmentaires
car ce sont des dérivés des somites et ce qui définit le niveau segmentaire c’est le niveau du
somite. Hors les sclérotomes structure segmentaire se divisent en 2 et se reconstituent pour
former les ébauches vertébrales inter segmentaires. La moitié caudal du sclérotome contribue
à la formation de la partie rostrale du corps vertébral et à l’ensemble de l’arc vertébral. La
moitié rostral du sclérotome contribue à la formation de la partie caudale du corps vertébral.
D’un point de vue finaliste, au niveau de chaque somite il y a émergence d’un nerf spinal, si
le nerf spinal sort du tube neural et bute contre le somite qui est juste en regard il ne va pas
pouvoir aller innerver les muscles moteurs dont il doit assurer l’innervation. Donc la nature a
retenu la solution de diviser le sclérotome en deux pour permettre au nerf spinal d’atteindre
les muscles dérivés qui sont juste un peu plus loin. Au niveau du tube neural, le bourgeon du
nerf spinal va droit devant, il n’y a aucun moyen de lui donner une information pour son
orientation donc le nerf spinal sort du tube neural, il trouve une cellule musculaire, il y adhère
ferme et ensuite tous les trajets nerveux sont liées qu’au fait que les cellules musculaires sont
dotées de capacités migratoire et le nerf va suivre. C’est aussi responsable d’un autre
problème d’anatomie : la partie rostral du 1er sclérotome cervical participe à la formation de
la base du crane et la partie caudal du 1er sclérotome cervical et les 7 sclérotomes cervicaux
suivant participent à la formation des 7 vertèbres cervicales ainsi qu’à la formation de la partie
rostral de la 1ère vertèbre thoracique. Au total on parle de 8 somites, 8 nerf spinaux pour 7
vertèbres cervicales. En conséquence les 7 premiers nerf cervicaux passent en avant des
vertèbres cervicales correspondantes, le 8ème nerf cervical passe entre la 7ème vertèbre cervical
et la 1ère vertèbre thoracique et à partir de là les nerfs spinaux suivant passent en arrière de la
vertèbre correspondante. Le disque intervertébral est disposé segmentairement donc chacun

3
est constitué de cellules conjonctives qui proviennent des sclérotomes et d’un noyau pulpeux :
nucléus pulposus constitué de cellules chordales qui disparaissent secondairement.
- cadran médio dorsal : forme le dermo myotome ou dermato myotome qui se divise
rapidement pour former le dermatome qui contribue à la formation du derme du tronc et au
myotome qui contribue à la formation des muscles axiaux du tronc. Les cellules les plus
dorsales du myotome forment l’épimère qui contribue à la formation des muscles rachidiens
et les cellules les plus ventrales forment l’hypomère qui contribue à la formation des muscles
des parois ventrale et latérale du thorax et de l’abdomen.
- moitié latérale : ces cellules de la moitié latérale migrent pour former les cellules
musculaires des membres.
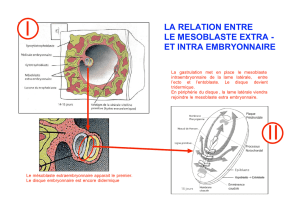
Délimitation embryonnaire
Elle commençait à la fin de S3 et elle se poursuit. Grossièrement on peut dire que la plicature
transversale de l’embryon amène les bords latéraux du disque embryonnaire à converger sur
la ligne médiane ventrale. Lorsque ces bords se rencontrent, les feuillet ecto, méso et
entoblastique (et amniotique) s’unissent à leurs homologues contro latéraux. A partir de
feuillets qui étaient plan on arrive à des cylindres.
Schéma :
A 20 jour, l’embryon est plan c’est le stade de la plaque neurale qui est en continuité avec
l’ectoblaste qui est lui même en continuité avec l’amnios de la vésicule amniotique. Le
mésoblaste : axial (chorde), para axial (somites), intermédiaire. Les lames latérales : la
somatopleure intraembryonnaire qui tapisse l’ectoblaste et qui est en continuité avec la
somatopleure extra embryonnaire qui tapisse l’amnios. La splanchnopleure intraembryonnaire
en continuité avec la splanchnopleure extra embryonnaire qui tapisse l’hypoblaste ou
entoblaste et la vésicule vitelline.
La plicature transversale se poursuit, il y a rapprochement vers la ligne médiane de
toutes les structures initialement latérales et au bout du compte il va y avoir fusion de
l’entoblaste gauche et droit, de la splanchnopleure gauche et droit, de la somatopleure gauche
et droite, de l’ectoblaste gauche avec le droit, fusion de l’amnios gauche avec l’amnios droit.
Et on arrive à cette configuration avec le tube neural, l’ectoblaste qui entoure complètement
l’embryon, l’intestin, le cœlome intraembryonnaire.
-Remarques :
-l’amnios contribue aussi à cette délimitation de l’embryon.
-ce processus de formation d’un cylindre ne va pouvoir être complet que dans la partie
rostral et caudal de l’embryon, il ne peut pas se poursuivre dans la région ombilicale sinon
l’embryon perdrait ses connexions avec ses annexes. On va traiter ce qui se passe au niveau
de la partie médiane de l’embryon c’est à dire la formation du cordon ombilical.
-au départ, on a pas de séparation clairement discernable entre le cœlome
intraembryonnaire et le coelome extra embryonnaire, ils communiquent à plein canal ; de
même qu’il est difficile de donner une limite précise à ce qui est la somatopleure intra et extra
embryonnaire ( et splanchno). A l’inverse quand les cylindres sont formés, le coelome
intraembryonnaire est très clairement isolé du coelome extra embryonnaire Il y a une phase
intermédiaire où souvent l’accolement temporaire de la somatopleure et de la splanchnopleure
contemporaine du processus de plicature transversale délimite le coelome intra embryonnaire
du coelome extra embryonnaire.
-Plicature sur une coupe longitudinale :
Ce qui a été décrit comme la formation des cylindres, c’est ce qui se passe dans les 10% des
les plus antérieur et les 10% les plus postérieur de l’embryon. A un stade plus avancé c’est ce
qui se passe au niveau des 30% les plus postérieur et les plus antérieur. Mais il y a un

4
problème : l’obstacle que constitue pour ce processus la vésicule ombilicale qui à ce stade est
encore très volumineuse, le pédicule embryonnaire qui contient l’allantoïde donc la fusion des
feuillets ectoblastiques conduit à ce que toute la surface embryonnaire soit recouverte
d’ectoblaste à l’exclusion d’une zone ventrale centrale où l’ectoblaste se réfléchit avec
l’amnios : c’est la région ombilicale où la vésicule vitelline et le pédicule embryonnaire
émerge. Et le développement de l’amnios au dépend du coelome externe est progressivement
responsable de la formation du cordon ombilical englobant le canal vitellin et allantoïde qui
finalement involue.
Le plexus vasculaire vitellin qui était siège de la vasculogénèse et de l’hématopoïèse primaire
va involuer lui aussi par contre le plexus vasculaire allantoïdien (celui qui s’est développé
autour de allantoïde) persiste et c’est à son niveau que se formeront les vaisseaux ombilicaux :
2artères et une veine.
-schéma : la cavité amniotique initialement c’était le 1/5 du volume du coelome externe mais
maintenant c’est le 1/3 de ce volume et déjà ça regroupe vers la partie ventrale de l’embryon
la vésicule ombilicale et allantoïde contenu dans le pédicule embryonnaire. Ce développement
de la cavité amniotique se poursuit au point que le coelome externe va devenir virtuelle (cours
suivant sur placenta). A ce stade le cordon ombilical est constitué : il englobe la vésicule
vitelline qui involue, allantoïde qui involue et on ne retrouve plus au niveau de ce cordon que
les 2 artères et une veine ombilicale. Ce cordon ombilicale est recouvert d’amnios et la zone
de réflexion à la face ventrale de l’embryon entre l’amnios et l’ectoblaste, c’est l’ombilic. A
28/30 J, la taille de l’ombilic est de 30% de la longueur totale de l’embryon (avant c’était
même 80%). Donc l’ombilic de 1cm représentait au départ 80% de la face ventrale de
l’embryon c’est à dire toute la zone où émergeait vésicule ombilicale et allantoïde
La fusion des lames latérales mésoblastiques entraîne la séparation définitive du coelome
interne et du coelome externe, cette fusion doit être envisagé distinctement selon qu’on se
situe en arrière ou en avant de la vésicule vitelline.
1- En arrière de la vésicule vitelline :
Le coelome interne devient la cavité péritonéale. La somatopleure fusionne, tapisse
intérieurement l’ectoblaste embryonnaire et constituera la séreuse péritonéale pariétale. La
splanchnopleure fusionne et tapisse extérieurement l’intestin, elle contribue à la formation du
tissu conjonctif, des feuillets musculaires du tube digestif et de sa séreuse péritonéale
viscérale. L’entoblaste embryonnaire est responsable de la formation de l’épithélium du tube
digestif mais tous les autres feuillets (conjonctif et musculaire) sont dérivés de la
splanchnopleure. Au départ l’intestin est largement relié à la partie dorsale du tronc par une
large bande de mésenchyme puis cette connexion s’amincit pour devenir un mésentère dorsal
constitué de l’accolement des lames splanchnopleurales. Cette cavité péritonéale à ce stade ne
contient rien, le tube digestif est rétro péritonéal. A un stade ultérieur lorsque le mésentère
s’est formé, le tube digestif le baigne pas librement à l’intérieur de la cavité péritonéale, il est
toujours recouvert par les dérivés de la splanchnopleure déjà il n’y a jamais rien dans la cavité
cœlomique interne sauf à chaque fois qu’il y a ovulation : l’ovule qui part de l’ovaire traverse
brièvement une partie du coelome interne avant d’être recueillit par la trompe utérine.

5
2- En avant de la vésicule vitelline :
- Mise en place des ébauches endothéliales (ébauche endocardique pour cœur)
Les cordons angioblastiques de la région cardiogène sont situés dans la moitié antérieure de
l’embryon autour de la plaque neurale, ils sont développés dans la splanchnopleure
intraembryonnaire et quand on regarde l’embryon de dessus on voit que ces cordons
angioblastiques sont autour de la plaque neurale. Ils confluent pour former les tubes
endocardiques latéraux qui sont fusionnés sur la ligne médiane en avant de la membrane
bucco pharyngée. Dans le même temps, il y a formation des aortes dorsales paire dans le
mésenchyme embryonnaire splanchnopleurale de part et d’autre de la ligne médiane. Et ces
aortes dorsales paire font leur jonction avec les tubes endocardiques au pole rostral de
l’embryon. Dans le même temps s’individualisent au pole caudal les afférences veineuses des
tubes endocardiques paire.
-Schéma : Au dessus d’un embryon à J20 il y a la vésicule vitelline qui à ce stade est
beaucoup plus large que le disque embryonnaire. Il y a la plaque neural et l’ectoblaste. Au
niveau de la vésicule vitelline il y a eu formation des plexus vitellins sièges de la
vasculogénèse et de l’hématopoïèse primaire mais au niveau de la splanchnopleure qu’on voit
à travers l’ectoblaste il y eu aussi formation de plexus vasculaires intra embryonnaire qui
confluent pour former les tubes endocardiques qui forment ce fer à cheval de part et d’autre
de la partie antérieur de la plaque neurale. Ces tubes endocardiques fusionnent sur la ligne
médiane en avant de la membrane bucco pharyngée qui est situé entre la partie antérieur de la
plaque neurale et la zone où fusionnent les tubes endocardiques. De la même façon par
transparence à travers la plaque neurale sont représentés les aortes dorsales paire qui
rejoignent les tubes endocardiques dans leur partie antérieur en avant de la membrane bucco
pharyngée. L’essentiel du cœur se développe en avant de ce fer à cheval, en avant de la
membrane bucco pharyngée.
En coupe sagittale il y a la plaque neurale en continuité avec l’ectoblaste (tapissé par la
somatopleure intraembryonnaire) puis en continuité avec l’amnios (tapissé par somatopleure
extra embryonnaire). L’entoblaste est en continuité avec l’entoblaste extraembryonaire.
L’accolement de l’ectoblaste et de l’entoblaste correspond à la membrane bucco pharyngée.
D’arrière en avant il y a la plaque neurale, la membrane bucco pharyngée puis ce qui va
former le cœur. Encore plus en avant cet accolement de la somatopleure et de la
splanchnopleure c’est le septum transversum qui est à l’origine de la formation du
diaphragme. Le cœur à ce stade est complètement au dessus de la tête et le diaphragme est
encore plus loin donc il y a un processus de réintégration du diaphragme et du cœur dans sa
position définitive et c’est ce qu’on va décrire.
- Description de la fusion sur la ligne ventrale des différents feuillets embryonnaires en
avant de la vésicule vitelline.
La fusion médiane de la splanchnopleure et de la somatopleure embryonnaire est responsable
de la séparation du coelome intra embryonnaire et du coelome extra embryonnaire et le
coelome intra embryonnaire devient la cavité pleuro péricardique primitive. Cette fusion est
responsable de la formation du tube cardiaque primitif qui est constitué par le tube
endocardique, la splanchnopleure qui est à l’origine de la formation du myocarde et d’un
manchon abondant de matrice extra cellulaire sécrété par la splanchnopleure et l’endocarde :
c’est la gelée cardiaque. La séparation du coelome interne et du coelome externe début sur la
ligne médiane et antérieur puis s’étends progressivement vers la partie moyenne de
l’embryon. Elle est responsable de la délimitation du septum transversum qui est un cordon
épaissit de mésoblaste dû à l’accolement de la somatopleure et de la splanchnopleure qui est
situé entre l’air cardiaque et le bord antérieur du disque embryonnaire. La délimitation
 6
7
6
7
1
/
7
100%