Cours du 05-02

Cours du 05/02/2008. 17h30 – 19h00.
Pr. Sterkers
REPONSE LYMPHOCYTAIRE T
PLAN
1- Généralités
2- Lymphocytes T naïfs
2-1 Répertoire
2-2 Migration programmée
2-3 Phénotype
2-4 Etude des lymphocytes circulants chez l’Homme
3- Présentation de l’antigène par les cellules dendritiques
3-1 Généralités
3-2 Dynamique
3-3 Synapse immunologique
4- Activation des lymphocytes
4-1 1er signal : intéraction TcR/MHC-Ag
4-2 2eme signal : intéraction CD28/CD80-86
4-3 3eme signal : rôle de l’Il2
4-4 Compétition entre signaux positifs et négatifs pour
l’acquisition d’une fonction
5- Prolifération des lymphocytes T
5-1 Expansion
5-2 Contraction

6- Différenciation fonctionnelle des lymphocytes T
6-1 T CD4 Th1
6-2 T CD8 cytotoxiques
6-3 Rôle des T CD4 pour l’acquisition d’une fonction cytotoxique
6-4 Modifications phénotypiques
7- Mécanismes de lyse
7-1 Sécretion des granules
7-2 Contenu des granules
7-2.1 Perforine
7-2.2 Granzyme B
7-2.3 Granzyme A
7-2.4 Granulosine
7-3 Déficits en molécules de mort
7-3.1 Déficit en perforine
7-3.2 Déficit en granzyme B
7-3.3 Déficit en granzyme A
7-3.4 Déficit en granzyme A et B
7-3.5 Déficit en cathepsine
7-4 Lyse dépendante de Fas
7-4.1 Voie d’activation de Fas
7-4.2 Déficit de la voie Fas / FasL
7-5 Immunopathologie
7-5.1 Hépatite fulminante
7-5.2 Chorioméningite lymphocytaire
7-6 Immunothérapie
8- Exploration lymphocytaire T
Je vous ai exposé le plan dans son intégralité, mais la prof n’a pas
tout traité. J’ai retranscris mot à mot son cours.

Nous allons traiter de la dynamique de la réponse immunitaire cytotoxique.
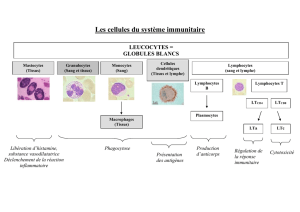
1- Généralités sur les CTL
Appartenance à l’immunité cellulaire
Il existe deux façons d’aider les hôtes immunoincompétents à se défendre contre les
agents infectieux.
L’immunité humorale : transférable par le sérum et due aux Ig.
L’immunité cellulaire transférable par des cellules.
(transfert d’une protection, à un individu naïf ou immuno-incompétent, par des cellules
immunes)
L’immunité cellulaire est une immunité déclenchée par la reconnaissance de l’antigène, elle
intervient en 2e plan après l’immunité naturelle. Les cellules spécifiques de l’antigène vont
donc proliférer et augmenter ainsi l’efficacité du système notamment par la
différenciation.
Il existe 2 types de réponses effectrices de l’immunité cellulaire :
- L’hypersensibilité retardée qui dépend pour son déclenchement des LT de
type Th1 (qui sécrètent de l’interféron ɣ). Exemple : les réponses
immunitaires contre les bactéries, en particulier les réponses immunitaires
contre le BK et l’exploration de cette immunité cellulaire, c’est
l’intradermoréaction.
- L’immunité déclenchée par les LTCD8 cytotoxiques et c’est celle là dont on
va parler aujourd’hui.
2- Lymphocytes T-CD8 naïfs
Définition : lymphocyte T-CD8 n’ayant pas encore rencontré l’Ag dont il est spécifique

2-1 Répertoire pré-immun
- diversité du répertoire (évaluée par la technique de immunoscope) 2 x 106
T-CD8. Pour pouvoir déclencher des réponses immunitaires efficaces, il faut qu’ils
soient capables de reconnaître les antigènes exogènes de la nature. Cette diversité est
réalisée dans le thymus où les thymocytes vont se différencier en LT. Au cours de
cette différenciation, il va y avoir des réarrangements des récepteurs T selon des
mécanismes qui seront générer par des recombinaisons VDJ, conduisant à une diversité
de l’ordre de 1015 spécificités. Après une double sélection dans le thymus pour éliminer
les clones T susceptibles de reconnaître des Ag du soi, si bien que le nombre de clones
T résiduels est de l’ordre de 108 à 1012.
Les récepteurs T vont reconnaître des fragments peptidiques de 9 à 15 résidus
présentés par des molécules du CMH. Il manque quelques clones, c’est ce que l’on appelle
des trous de répertoire mais globalement cette diversité permet au système
immunitaire de reconnaître la quasi-totalité des Ag exogènes.
Donc ces LT matures après cette recombinaison et cette double sélection vont sortir du
thymus et constituent le répertoire T naïf très divers.
Ces LT naïfs vont avoir une seule chose à faire initialement c’est à essayer de rencontre
l’Ag dont il est spécifique. Pour cela, il existe un système de migration programmée qui est
assez efficace et qui permet aux LT naïfs de reconnaître les Ag exclusivement dans les
ganglions qui limitent l’espace dans lequel les LT vont pouvoir rencontrer leurs Ag.
2-2 Migration programmée
Les lymphocytes naïfs émergent du thymus et passent dans la circulation, et dans la
circulation et migrent de ganglions en ganglions dans l’organisme.
A chaque fois qu’ils arrivent dans un ganglion, l’Ag spécifique du lymphocyte sera dans le
ganglion, et sera donc reconnu et se différencie en L mémoire qui eux changent de
phénotype, et au lieu de migrer de ganglion en ganglion, ils vont rester vers les tissus et en

particulier le tissu infecté à ce moment là. Ils jouent donc leurs rôles de lymphocytes
effecteurs dans le tissu infecté.
Comment rencontrent- ils l’Ag dans le ganglion ?
C’est parce que les CPA en réponses primaires sont les cellules dendritiques, qui ont une
migration programmée inverse des LT naïfs, à savoir qu’elles résident dans le tissu, et que
lors d’infection, non seulement ces cellules dendritiques vont capter l’Ag, mais
l’inflammation locale va leur donner un signal de migration vers le ganglion drainant où se
retrouvent alors les Ag. Les CPA une fois qu’elles ont présenté l’Ag meurent dans le tissu
infecté.
- permet la rencontre entre le lymphocyte et l’antigène dont il est spécifique.
- cette migration est inversée par rapport aux cellules dendritiques qui sont dans les
tissus et migrent vers les ganglions en présence de l’antigène
Migration programmée
Cellules dendritiques
chargées d’Ag
lymphocytes mémoires
1.1.1.1.1.1.1
2-3 Phénotype :
1.1.1.1.2 A
g+
A
g-
1.1.1.1.2.1.1.1 Lym
phoc
ytes
naïfs
1.1.1.1.1.1.1.3 C
i
r
c
u
l
a
t
i
o
n
1.1.1.1.1.1.1.1 G
a
n
g
l
i
o
n
1.1.1.1.1.1.1.2 T
i
s
s
u
s
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
1
/
25
100%