ONTOGENESE VEGETALE

Transition
florale
fécondation
Gamètes n
Sénescence
programmée et mort
Stade reproducteur
Plante mature
stade végétatif
plantule
Embryon
(graine)
Zygote 2n
germination
croissance
ONTOGENESE VEGETALE
1. Cycle de développement et reproduction des plantes
2. Grandes lignes de la formation de l’appareil végétatif
3. Racines et méristème racinaire
4. Tige et méristème apical
5. Transition florale et inflorescence
6. Fleurs
7. Gamétophytes et gamètes
8. Stratégies de reproduction, cycle de développement des angiospermes.
Une plante est fixée sur le sol. Elle doit donc s’adapter aux changements de son environnement et aux
attaques biotiques. C’est pourquoi l’ontogenèse et le développement répondent à des mécanismes d’une grande
plasticité, selon les conditions du milieu.
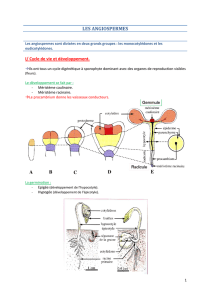
Le cycle de développement d’un individu :
Chacune de ces étapes est dépendante des conditions du milieu.
La plantule est un individu encore hétérotrophe, c’est à dire qu’il n’a pas encore différencié son appareil
photosynthétique. Le stade végétatif correspond à un stade où aucun organe reproducteur n’est exprimé. A ce
stade, de nombreux organes sont produits : feuilles, tiges, racines, bourgeons. Les organes reproducteurs sont
néoformés : ils ne se forment pas dès le stade embryonnaire. Il n’y a pas de lignées germinales. A certains stades
(graine, bourgeon), il y a possibilité de vie ralentie ou dormance. C’est un programme de protection contre le
stress thermique, hydrique, accumulation de réserves…sous contrôle hormonal.
Quelques notions de taxonomie :
L’ensemble des êtres vivants est divisé en deux groupes, les eucaryotes et les procaryotes. Les procaryotes sont
formés par les archéobactéries et les eubactéries, les eucaryotes par les animaux, végétaux et champignons. On
désigne par le monde végétal l’ensemble des organismes capables de photosynthèse. Il existe plusieurs groupes
qui suivent l’évolution au cours des temps géologiques :
Thallophytes : algues : l’appareil végétatif est simplifié à des couches cellulaires peu organisées (thalle).
Cormophytes : végétaux à tige, cela est lié à la colonisation du milieu terrestre.
- Bryophytes : mousses, sphaignes. Pas d’enracinement profond, pas de vaisseaux.
- Brachiophytes : végétaux à vaisseaux et racines.
- Ptéridophytes : fougères, sélaginelles…
- Phanérogames :
- Gymnospermes : conifères ; graines sur l’extérieur
- Angiospermes mono et dicotylédones : plantes à fleur,
graines dans un fruit.
Cycle de reproduction :
Un végétal passe par deux stades distincts, soit deux individus, l’un diploïde l’autre haploïde, le sporophyte et le
gamétophyte. On parle de cycle de reproduction digénétique. Un cycle digénétique peut être isomorphe ou non,
c’est à dire qu’une forme haploïde ou diploïde peut être favorisée. Si les sexes sont séparés, on parle d’individus
dioïques. Il existe aussi des cycles monogénétiques haploïde où le zygote subit très vite la méiose. La forme
sporophyte est en fait le zygote. Mais la diploïdie ne permet pas une grande multitude des allèles donc une

Gamètes n
Méiose n
Sporophyte
Zygote 2n
Gamétophyte
Méristème
racinaire
Zone de
différenciation
Méristème apical
sol
Zone d’élongation
grande souplesse d’adaptation. Ainsi les cycles monogéniques diploïdes sont favorisés. Les spores constituent le
gamétophyte. Ce sont très souvent des cycles hétéromorphes.
Il peut aussi exister des cercles trigénétiques (algues rouges) où il y a doublement de la génération diploïde. Les
bryophytes classiques sont gamétophytes. Les sporophytes se développent de façon parasitaire sur le
gamétophyte. Les trachéophytes ont un cycle digénétique hétéromorphe où le sporophyte est dominant. Le
pollen est le gamète mâle. L’ovule est le gamétophyte femelle.
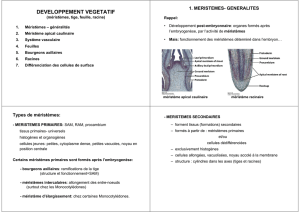
L’appareil végétatif se construit en permanence au cours de la vie de la plante. Les méristèmes sont responsables
de cela. Ce sont des zones de cellules douées d’activité mitotique organisées (contrairement à une tumeur). Il
s’en suit une histogenèse et une organogenèse. Ces méristèmes sont localisés spécifiquement aux extrémités :
méristème apical responsable de l’édification de la tige principale, bourgeon axillaire pour l’édification des
nouvelles branches. Il existe des phénomènes de dominance entre méristèmes apicaux et latéraux. Le système
racinaire est formé par un méristème apical racinaire mais il n’existe pas de méristèmes latéraux pour les racines.
Les méristèmes secondaires sont néoformés. Il existe aussi des cambium responsables de la croissance en
diamètre. Mais un méristème conduit à de petites cellules. Il faut une étape de croissance cellulaire. Ce
phénomène est localisé dans les tissus en dessous des méristèmes, dans les parties jeunes. Le grandissement
cellulaire nécessite de l’eau (entrée par diffusion dans la cellule, selon le potentiel hydrique). Cela est contrôlé
par la teneur en ion intracellulaire. La paroi cellulosique doit rester déformable (plasticité) sous la pression de
turgescence, avec une notion de seuil.
Il faut noter quelques originalités des plantes sur les animaux :
- Organogenèse permanente liée aux méristèmes
- Pas de migration cellulaire à cause de la paroi pectocellulosique et absence de cellules souches (lignées
cellulaires). L’ontogenèse des plantes est due essentiellement à l’effet position. Cet effet position est dû à un
ensemble de gradients trophiques (ressources), hormonaux, électriques, de pression…
Il existe aussi un contrôle de l’activité mitotique, qui est organisée dans le méristème, comme un contrôle du
plan de division : des divisions anticlinales vont élargir le tapis de cellules tandis que des divisions périclinales
vont provoquer une percée du massif de cellules. Un autre contrôle se fait au niveau de l’intensité et de
l’orientation du grandissement cellulaire (croissance isodiamétrique ou polarisée), mais aussi l’identité des
organes (gènes homéotiques).
Chez le végétal, la cellule indifférenciée est appelée cellule méristématique, ou cellule embryonnaire. Ce sont
des cellules qui ont une activité mitotique organisée importante et qui n’ont pas de différenciation tissulaire
particulière. Ces cellules évolueront en cellules différentes (pas de lignées cellulaires). A l’intérieur d’un organe,
la cellule différenciée ne se divise plus mais elle conserve sa potentialité de dédifférenciation pour s’engager
vers une autre voie de différenciation. Par exemple si on lèse les vaisseaux dans une tige, on observe que

Gemmule : tige embryonnaire et méristème apical
caulinaire
Cotylédones
Radicule
Epiderme :
Trichoblastes ou
Atrichoblastes
Zone pilifère
certaines cellules du parenchyme vont se re-différencier en cellules conductrices, pour créer une circulation
alterne. C’est la réversibilité des étapes de différenciation. On utilise cette capacité pour le bouturage. C’est la
totipotence des cellules végétales. Les plantes peuvent aussi avoir la capacité de néoformer des organes
spécifiques par l’intermédiaire de bactéries. C’est le cas chez les Légumineuses qui en présence de Rhizobium
vont former des nodosités pour les accueillir, à partir de cellules du cortex racinaire, pour une symbiose autour
de N2 du sol. C’est le cas aussi des galles, souvent liées à des pathogènes (œufs d’insectes ou bactéries) qui
libèrent des hormones et induisent la différenciation des cellules du végétal.
La culture in vitro utilise la totipotence des cellules végétales : à partir d’un explant, on peut réaliser des cultures
de cellules relativement indifférenciées en milieu liquide (suspension) ou solide (cal), des cultures de
protoplastes (paroi digérée) ou culture de fragment d’organes ou de tissus. On peut diriger les différenciations
par des hormones. On travaille beaucoup sur une plante modèle Arabidopsis thaliana (http://genome-
www.stanford.edu/arabidopsis). Elle est avantageuse du point de vue de la taille, elle est autogame
(homozygotie), cycle court, mutagène, fine et relativement transparente.
Racine et méristème racinaire :
La racine est un organe très différent des parties aériennes car elle ne connaît pas la transition florale. Elle a un
développement continu issu du fonctionnement du méristème apical racinaire MAR. La racine a plusieurs
fonctions : absorption d’eau et ions minéraux, ancrage, stockage, conduction. Elle est présente dès l’état
embryonnaire
La radicule évolue en racine primaire (axe principal). Elle peut ensuite se ramifier en plusieurs racines latérales
ou secondaires (cf. TD UE3)
Les tiges se ramifient selon une base répétées, le métamère qui porte une feuille et un bourgeon. Il n’y a pas
d’équivalent chez la racine. Elle apparaît à ce sujet un peu plus chaotique.
Le primordium (massif de cellules qui va former la nouvelle racine) est le résultat d’une dédifférenciation de
cellules de la racine primaire et non de MAR.
Les racines ont un gravitropisme positif et un phototropisme négatif.
Elles possèdent une structure très particulière, les poils absorbants. Ce sont des expansions de cellules de
l’épiderme
La zone pilifère se déplace en suivant la croissance de la racine.
Organisation du MAR :
On observe des fils de cellules qui forment les différents tissus.
Métamère
Racine primaire
Racine
secondaire
Cylindre central :
- endoderme subérifié (interrompt la
voie apoplastique intracellulaire de
diffusion des substances
- péricycle à partir duquel se
forment les racines secondaires.
- Faisceau de xylème et phloëme
- Parenchyme central.
Epiderme
Parenchyme
cortical

Stèle
Cortex+endoderme
Columelle
latérale+épiderme
Cellules quiescentes
Columelle centrale
méristème apical+cotylédones
hypocotyle+racine
hypophysecoiffe+centre quiescent
Divisions
asymétrique
s
zygote
Arabidopsis thaliana a une organisation de ces tissus très simple, sur seulement une assise cellulaire.
Le MAR est constitué de 2 types cellulaires :
les initiales :
- initiales de la columelle centrale (divisions périclinales)
- initiales de la columelle latérale et épiderme (anticlinales puis périclinales)
- initiales du cortex et endoderme (anticlinales puis périclinales)
- initiales de la stèle (périclinales)
les cellules quiescentes qui se divisent peu ou pas du tout
Localisation des initiales dans le MAR :
CL :
Au-dessus de la zone de division méristématique, on trouve la zone d’allongement cellulaire puis la zone de
différenciation.
Pour identifier les zones mitotiques, on peut colorer les noyaux pour observer les figures de mitose.
Les cellules quiescentes entretiennent le caractère de développement infini du MAR.
La zone d’allongement est très importante pour le grandissement polarisé des cellules.
Le MAR est formé chez l’embryon, avec ses initiales :
Le MAR est donc issu de l’hypophyse de l’embryon et de la demi-partie supérieure. Mais ces frontières restent
floues. On dispose de marqueurs génétiques pour suivre la mise en place des initiales, par exemple le gène GUS
(glucuronidase). Cette enzyme est en effet repérable grâce à un substrat chromogène. Le gène GUS est inséré
dans le génome de la plante avec des promoteurs spécifiques des territoires cellulaires coiffe, columelle… une
fois le territoire cellulaire repéré, on détruit précocement ce qu’on pense être une initiale et on observe
l’influence sur le développement. On réalise des ablations cellulaires, ou destruction par laser. On met ainsi
l’importance de l’effet position : en détruisant une initiale, on observe qu’une autre cellule prend sa place et
remplit son rôleil n’y a pas de cellules souches mais un effet position.
Mais pour qu’une cellule acquière un statut précis (comme celui d’initiale), elle doit être en contact avec des
cellules différenciées. C’est une cellule différenciée qui va reprogrammer une cellule en initiale. Cela a été
prouvé par des expériences d’ablation. C’est la communication intercellulaire (composition chimique de la paroi
Coiffe
(cap)
Columelle
centrale
Columelle
latérale
MAR
Stèle
Parenchyme cortical

SAM
Primordium foliaire
Feuille
L1 : divisions anticlinales
L2 : divisions anticlinales et
quelques périclinales
L3 : anti et périclinales
(centrestèle)
CZ : central zone : peu de mitoses
PZ : peripherical : activité mitotique
plus importante
RibZ : médullairetissus de la stèle.
3
2
1
et par les plasmodesmes). Ces mécanismes de communication sont peu connus. On possède des mutants pattern
qui modifient leur organisation générale, des mutants d’activité méristématique (prolifération ou arrêt précoce)…
Le méristème apical caulinaire SAM (shoot apical meristem) :
Il a pour fonction de fabriquer toutes les cellules de la tige. Il est localisé à l’extrémité de la tige.
Le SAM établit la succession des métamères de la tige : nœud, feuille, bourgeon axillaire.
Le bourgeon axillaire, en dormance au pied de la feuille permettra la ramification. La plante acquière ainsi une
architecture aérienne particulière, dépendante de sa programmation génétique et des conditions du milieu. Le
SAM est formé dès le stade embryonnaire. Il produit tous les tissus végétatifs et, grâce à un changement
d’activité, les pièces reproductives (méristème d’inflorescence). Le SAM caractérise le méristème végétatif.
Structure du SAM :
Il y a plusieurs façons de caractériser les différents éléments du SAM : par couche cellulaire (layers) ou par
zones :
Il y a beaucoup de variantes à cette organisation : les mousses et fougères ne possèdent qu’une seule cellule qui
sert d’initiale à toute la tige, les monocotylédones et gymnospermes ne possèdent pas de L2…
Pour étudier la formation et l’entretien du SAM, on peut réaliser des mutations dirigées qui affectent pare
exemple la teneur en chlorophylle ce qui est repérable par la suite. On obtient une plante chimère ou mosaïque,
car toutes les cellules n’ont pas le même génotype. On a pu montrer ainsi que L1 forme l’épiderme, L2 le bord
du limbe des feuilles et une partie des tissus périphériques de la tige et L3 la majeure partie de la tige, avec une
grande variabilité. Dans chaque couche il y a de 1 à 3 cellules à l’origine de toutes les autres cellules. On peut les
qualifier d’initiales. ces initiales sont à l’origine de secteurs de la plante. Mais ces frontières ne sont pas fixes, il
existe des échanges de cellules.
Il n’y a pas de lignées cellulaires fixes, là encore c’est l’effet position qui est prépondérant. Il n’y a pas de
lignage génotypique.
Les initiales se situent au sommet de la zone centrale. Dans la PZ, on a déjà un début de différenciation. La zone
centrale apparaît conférer au Sam un caractère de développement infini.
Pour étudier le fonctionnement u SAM, on réalise sur des mutants des travaux de profil d’expression du gène,
avec des hybridations in situ. On appelle caulogenèse la fabrication de la tige, qui est le résultat du
fonctionnement du SAM.
Les méristèmes sont des structures relativement indépendants : si on décapite le SAM et qu’on le place sur un
milieu de culture adéquat, il continue de fonctionner de façon indépendante pour produire une plantule entière.
Cette propriété est utilisée pour sauver une plante de l’attaque d’un virus. Comme le virus n’attaque jamais le
SAM, on peut ainsi reconstruire une plante saine entière. Cela permet aussi une culture in vitro intensive.
La phyllotaxie est aussi utilisée pour déduire le fonctionnement du SAM : c’est l’étude de la disposition des
feuilles le long de la tige, qui rend compte de l’activité du SAM. On déduit ainsi par des expérience de dissection
des primordium foliaires qu’un primordium inhibe la formation d’un autre primordium trop près :
Cet effet inhibiteur de l’organe déjà formé est assez général pour d’autres organes.
Les méristèmes sont aussi capables de néoformation, à partir de cellules différenciées ou indifférenciées, sur une
tige qui n’a pas de SAM. Il se forme par recrutement de plusieurs cellules en fonction de leur position et de
signaux hormonaux.
 6
7
8
9
10
11
12
13
14
15
6
7
8
9
10
11
12
13
14
15
1
/
15
100%