La reproduction sexuée des organismes supérieurs

I) Introduction
La reproduction sexuée des organismes supérieurs
Les différents types de reproduction :
- asexuée chez un organisme unicellulaire : la mitose, les cellules filles contiennent un matériel génétique
identique à celui de la cellule mère (ex : amibe).
- bourgeonnement chez des organismes pluricellulaires : les cellules du bourgeon viennent de mitoses répétées
de cellules parentales. Le bourgeon est génétiquement (presque) identique à l’organisme parental, les rares
différences sont dues à des mutations par des agents mutagènes (ex : hydre).
- division et régénération d’organismes pluricellulaires : ils sont scindés en 2 et chaque demi-organisme
régénère la partie manquante (ex : étoile de mer, vers marins).
Ces reproductions asexuées donnent naissance à un clone.
La reproduction sexuée, elle, implique un mélange de deux génomes de deux individus différents :
- elle donne naissance à des individus contenant chacun une combinaison unique de gènes
- le mélange est obtenu par fusion des cellules haploïdes
- elle nécessite une réduction de la ploïdie de moitié pour permettre un nombre constant de chromosomes de
l’espèce après la fusion.
Ploïdie = nombre de jeux de chromosomes par cellule, n 2n 3n 4n….
Méiose = division cellulaire impliquant une réduction de la quantité de matériel génétique (grec meios =
réduction).
Gamètes = cellules haploïdes spécialisées dans la fusion sexuée
Fécondation (syngamie) = union de deux gamètes de sexe différent
Zygote = cellule diploïde, produit de la fécondation
Cycle de développement = suite des étapes qui se déroulent depuis la conception d’un organisme jusqu’à
l’apparition de ses propres enfants
Cycle de reproduction = alternance de phases haploïdes/diploïdes

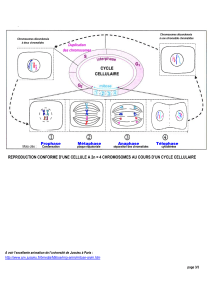
II) La méiose
Elle est composée de 2 divisions successives. De S à G2 on a une duplication de l’ADN par réplication amenant
à 2n chromosomes à 2 chromatides, la phase G2 est courte. La méiose I est une division réductionnelle où on
passe de 2n à n chromosomes, les chromosomes homologues sont séparés en deux lots. La méiose II est
équationnelle, elle consiste en la séparation des chromatides. On obtient 4 cellules haploïdes à 1 chromatide
appelées gamètes. Il existe un échange de matériel génétique entre chromosomes homologues, la recombinaison,
permettant un brassage génétique.
A) Méiose I
1) Prophase I : reconnaissance réciproque puis appariement des chromosomes homologues à 2 chromatides grâce
à des mécanismes de recombinaison (échanges de segments d’ADN). Ces recombinaisons ne peuvent avoir lieu
qu’entre séquences nucléotidiques partiellement identiques. La prophase I peut occuper 90% de la durée totale
de la méiose. Elle comprend 5 stades :
- leptotène : le noyau grossit, les chromosomes deviennent apparents sous forme de pelote enchevêtrée de
filaments longs et minces. Un ou plusieurs nucléoles sont présents. Un assemblage de protéines le long des
chromosomes se produit.
- zygotène : les chromosomes se condensent et deviennent plus courts, les télomères visibles sont ancrés à la
lamina nucléaire, les chromosomes homologues commencent à s’apparier en formant des bivalents : ce processus
est appelé synapsis. L’assemblage d’une structure protéique : le complexe synaptonémal, entre les
chromosomes homologues, apparaît. On ne sait pas encore comment deux chromosomes homologues se
rencontrent, mais on pense que les télomères se trouvent à proximité, ancrés par la lamina.
Les 2 chromatides sœurs de chacun des chromosomes sont étroitement associées et se déploient du même côté de
l’échelle protéique. Entre 2 éléments latéraux, on a une distance de 200 nm (ce qui est énorme). Les
chromosomes ne pouvant pas s’apparier ne peuvent pas recombiner et la méiose n’est pas possible. Des mutants
capables de recombinaison sans complexe synaptonémal (et donc capables de faire une méiose) ont cependant
été trouvés, mais une recombinaison est nécessaire pour la méiose. Le rapprochement des chromosomes avec des
recombinaisons aurait lieu très tôt dans la prophase, et le complexe synaptonémal ne se formerait qu’après.

- pachytène : (pachos, grec = épais) les chromosomes sont courts et épais, l’appariement s’achève, le complexe
synaptonémal est entièrement mis en place. Les enjambements chromosomiques ont lieu, ils impliquent une
cassure et une réunion croisée très précise des doubles hélices d’ADN de deux chromatides homologues non
sœurs. Les nucléoles sont souvent bien définis. Ce stade peut durer plusieurs jours. Les nodules de
recombinaison permettent les enjambements, ce sont des complexes multi-enzymatiques. On pense que la
formation du complexe synaptonémal résulte de l’initiation de la recombinaison.
- diplotène (diplos, grec = double) : le complexe synaptonémal se désintègre et les homologues s’écartent,
révélant des structures cruciformes : les chiasmas (manifestation visible des enjambements). C’est une période
de croissance extrêmement longue pendant laquelle les chromosomes se décondensent et ont une transcription
très active. Les ovocytes humains sont en stade diplotène du 5ème mois fœtal à l’ovulation qui débute à la puberté
et s’arrête à la ménopause.
- diacinèse : les chromosomes se condensent à nouveau et se détachent de l’enveloppe nucléaire. Les chiasmas
migrent vers les extrémités des chromosomes, ce processus est appelé terminalisation. Les deux asters ont migré
aux pôles opposés du noyau et le fuseau de division s’assemble. Les nucléoles disparaissent. Les bivalents
s’écartent par un processus appelé asynapsis et la distinction des chromatides sœurs devient plus évidente. Le
nombre de chiasmas par chromosome est variable (2 à 3 enjambements ou crossing overs par paire de
chromosomes, il n’y en a pas au voisinage de l’hétérochromatine). La fréquence de recombinaison est 2 à 3 fois
plus élevée chez les femmes que chez les hommes.
Ne pas confondre enjambement (superposition des chromatides non sœurs homologues avec échange de matériel
génétique par cassure) et chiasma (conséquence de l’enjambement, croisement des chromatides visible et qui se
déplace vers les télomères, alors que la position de l’enjambement reste sur place).
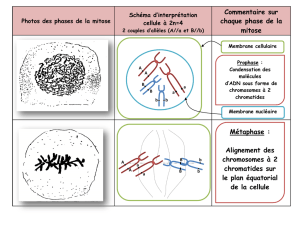
2) Métaphase 1 : il n’y a qu’un seul kinétochore par centromère. La condensation est maximale, les microtubules
capturent les bivalents et les placent sur la plaque équatoriale. La protéine Mam I entraîne la fusion des
kinétochores des chromatides sœurs, et par conséquent les kinétochores des chromosomes homologues sont
attachés aux pôles opposés du fuseau. Ils sont toujours liés l’un à l’autre par des chiasmas. Sans recombinaison,
la méiose ne peut pas avoir lieu.
3) Anaphase 1 : initiée par une dissolution simultanée des chiasmas, les homologues migrent vers les deux pôles
opposés du fuseau. Leur répartition vers l’un ou l’autre pôle est laissée au hasard. La protéine Mam I protège les
cohésines contre un clivage par la séparase et par conséquent les chromatides sœurs restent liées.
4) Télophase 1 : elle est brève, l’enveloppe nucléaire se referme pendant peu de temps. Le nombre de
chromosomes de chaque cellule fille a été réduit de moitié, ils possèdent 2 chromatides sœurs qui ne sont pas
identiques à cause de la recombinaison.
Entre les deux divisions méiotiques, une intercinèse a lieu : c’est une courte interphase sans synthèse d’ADN.
B) Méiose II
1) Prophase 2 : elle est très courte, l’enveloppe nucléaire disparaît, le fuseau de division se forme et les
chromosomes se condensent.
2) Métaphase 2 : les chromatides sœurs possèdent maintenant chacune un kinétochore lié à un jeu de
microtubules venant des pôles opposés, elles s’écartent l’une de l’autre (cohésines détruites à la fin de la
métaphase 1 : la Mam I n’a protégé que les cohésines se trouvant à proximité du centromère, les autres ont été
détruites).
3) Anaphase 2 : la protéine Mam I va être dégradée ce qui permet la séparation des chromatides sœurs qui sont
tirées aux pôles opposés de la cellule.
4) Télophase 2 : l’enveloppe nucléaire se reforme autour de 4 lots d’ADN. De la cytocinèse résultent 4 cellules
haploïdes. Chaque noyau contient une combinaison unique de gènes due à la recombinaison pendant la prophase.
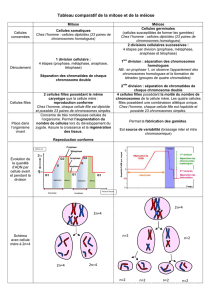
C) Comparaison mitose – méiose
Mitose
Méiose
1 seule division
2 divisions successives sans synthèse d’ADN à

l’intercinèse, échanges de fragments d’ADN en prophase 1
Séparation des chromatides sœurs uniquement, pas
de changement de ploïdie
Séparation des chromosomes homologues en méiose I
provoquant un changement de ploïdie
Production de 2 cellules diploïdes génétiquement
identiques entre elles et à la cellule-mère
Production de 4 cellules haploïdes qui possèdent chacune
une combinaison unique de gènes
Se produit dans toutes les cellules somatiques
Apparaît uniquement dans les cellules germinales localisées
dans les tissus spécialisés : les gonades
Responsable de la croissance de l’organisme et de la
réparation tissulaire en cas de lésion
Destinée exclusivement à la production de gamètes
D) Le brassage génétique
3 phénomènes contribuent à la diversité génétique de la reproduction sexuée :
- répartition au hasard des homologues maternels et paternels entre les cellules filles au cours de la division
méiotique I. Chaque individu peut produire 2n gamètes génétiquement différents (n = nombre haploïde de
chromosomes, soit 23 chez l’homme). 223 = 8,4.106 gamètes différents
- échanges génétiques permettant de combiner dans un même chromosome l’ADN hérité des deux parents
(augmentation du nombre réel de gamètes différents produits)
- nature aléatoire de la fécondation augmentant la variété génétique : homme 8,4.106 x 8,4.106 = 70,56.1012
combinaisons possibles
On peut aussi ajouter les mutations nucléotidiques qui causent elles aussi une diversité.
http://johnkyrk.com/meiosis.html
III) La gamétogenèse (voir schéma)
Les cellules germinales primordiales réalisent 2 mitoses puis on obtient :
Spermatozoïde primaire (4 Chromosomes à 2 Chromatides
(méiose I) Spermatozoïde secondaire 2 Chromosomes à 2 Chromatides
(méiose II) Spermatides Spermatozoïdes 2 Chromosomes à 1 Chromatide)
Ovocyte primaire (4 Chromosomes à 2 Chromatides
(méiose I) Ovocyte secondaire 2 Chromosomes à 2 Chromatides
(méiose II) Ovule 2 Chromosomes à 1 Chromatide)
Les spermatozoïdes sont petits et mobiles, les ovules sont volumineux (100 µm de diamètre), immobiles,
totipotents et hautement spécialisés.
A) Ovogenèse
1) Les divisions méiotiques

En absence d’hormones androgènes, les gonades primitives se transforment en ovaires à partir de la 9ème semaine
fœtale. Vers le 5ème mois, les cellules souches germinales (diploïdes) appelées ovogonies se multiplient par
mitoses. Pendant la dernière partie de la vie fœtale, les ovogonies se transforment en ovocytes primaires, elles
amorcent la méiose I et s’arrêtent à la prophase I (stade diplotène). La fille naît avec 700 000 ovocytes primaires
situés dans le follicule primaire. Avant la naissance, chaque ovocyte s’entoure d’une seule couche de cellules
nourrcières.
A partir de la puberté, par contrôle hormonal, la deuxième division méiotique est amorcée par vagues. Un seul
devient ovocyte II et est expulsé d’un follicule mature lors de l’ovulation. Celui-ci est bloqué en métaphase II,
entouré par la granulosa.
2) Maturation de l’ovocyte et du follicule
La taille de l’ovocyte I augmente suite à un signal hormonal, avec la synthèse de matériaux nécessaires en cas de
fécondation. L’ovocyte I s’entoure d’une couche de glycoprotéines, la membrane pellucide. Les cellules
folliculaires prolifèrent et donnent naissance à plusieurs couches.
Peu avant l’ovulation, l’ovocyte I termine sa méiose I. Le fuseau se forme à l’extrême bord de la cellule et tous
les organites quittent cette région. La cytocinèse est asymétrique, créant une petite cellule (1er globule polaire)
sans organites ni cytoplasme et une grosse cellule (ovocyte II) qui conserve le cytoplasme de la cellule mère.
Parallèlement à la croissance ovocytaire se déroule la croissance du follicule qui passe en 14 jours de 40 µm à 20
mm. Un follicule mature est constitué de plusieurs couches cellulaires, la thèque et la granulosa, qui délimitent
une cavité liquidienne, l’antrum. Cette cavité contient l’ovocyte arrêté en métaphase II.
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%