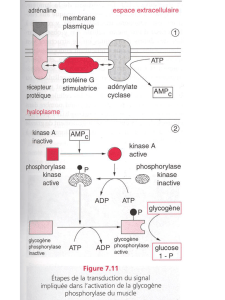
B - Les autres hormones

1/23
SOMMAIRE
Introduction ......................................................................................page 1
I - Facteurs Environnementaux ................................................... page 2
A – Généralités .........................................................................page
B - Les effets du climat ..........................................................page
1) Effets de la Température
2) La Pression de Turgescence
C - Les Tropismes .....................................................................page
1) Le Phototropisme
2) Le Géotropisme
3) Autres Tropismes
II – Action des Hormones sur la Croissance .......................................... page
des Végétaux
A - L’auxine ........................................................................................... page
B - Autres Hormones .......................................................................... page
C - Action simultanée des différentes Hormones ....................... page

2/23
Conclusion ........................................... pageIntroduction
L ‘ensemble du règne végétal comporte beaucoup d’espèces, même si leur nombre est
faible par rapport à celui du règne animal. Son importance au sein de la biosphère est
considérable : les végétaux sont capables de transformer la matière minérale en matière
organique. De ce fait, ils forment le premier maillon de toutes les chaînes alimentaires et
agissent sur la composition du milieu, en particulier de l’atmosphère. Ils sont le support de
toute vie sur Terre.
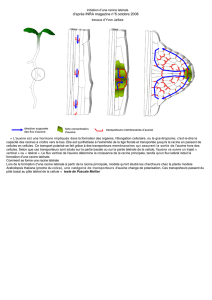
La morphogenèse d’un végétal est la réalisation de la structure d’embryons, d’organes, de
tissus chez les êtres vivants, afin que leur formation ne soit pas due au hasard. Les plantes
utilisent surtout l’énergie lumineuse, les animaux l’énergie chimique des substances que
fabriquent les plantes. Dans ce TPE nous nous intéressons uniquement à la morphogenèse des
végétaux, et plus précisément ici au légumineuses herbacées à valeur alimentaire. En effet,
dans notre choix des plantes à sélectionner pour étudier leur développement et les différents
facteurs pouvant le modifier, nous avions besoin de plantes qui poussent relativement vite,
que nous pourrions mettre en culture en grande quantité, et qui ne seraient pas très complexe à
faire pousser.
Nous nous sommes donc demandées quels étaient les différents facteurs pouvant modifier la
morphogenèse d’un végétal. Nous développerons à la fois les facteurs externes
(environnementaux), mais aussi les modifications intracellulaires engendrées par ces facteurs
qui peuvent avoir une influence sur leur morphogenèse.I – L’influence de
l'environnement
Quand l’origine du signal qui déclenche un facteur morphogénétique est en dehors de
l’organisme, c’est l’environnement, grâce aux variations d’un ou de plusieurs de ses
composants physique(lumière, température) ou chimique(substances de croissance, éléments
de la nutrition minérale), qui va indirectement déterminer la réponse morphogénétique. Nous
n’avons pas pris en compte tous les facteurs pouvant modifier la croissance, car cela aurait été
trop lourd. Comment l’environnement va contribuer à modifier la croissance ?Quels sont les
mécanismes intracellulaires mis en jeu ?
A - Généralités
Quels sont les mécanismes intracellulaires qui permettent la croissance d’une cellule
végétale ?
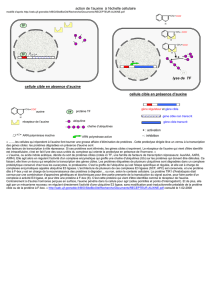
La cellule végétale
Comme la cellule animale, la cellule végétale comprend un cytoplasme renferment un
noyau et divers organites. Mais ce cytoplasme ne constitue qu’une mince couche
périphérique, car il comporte une vacuole centrale. La cellule dispose d’une paroi qui est une
coque rigide indépendante. La cellule contient des organites : les mitochondries qui jouent un
rôle dans les phénomènes d’oxydation et de stockage de l’énergie, les chloroplastes qui
contiennent de la chlorophylle pour la photosynthèse.

3/23
Le développement des plantes à graines
Le développement d’un organisme, c’est-à-dire son évolution au cours du temps, se
traduit extérieurement par deux séries de modifications :
des modifications quantitatives qui se traduisent par une augmentation des dimensions,
comme la surface, la longueur, le volume ou la masse. C’est la croissance.
des modifications qualitatives, qui se traduisent par l’acquisition de propriétés nouvelles,
morphologiques et fonctionnelles. C’est la différenciation.
La Croissance
Dans le cas d’un organisme pluricellulaire, la croissance peut être obtenue soit par
croissance des cellules déjà existantes, soit par augmentation du nombre de ces cellules. Il y a
trois processus :
la prolifération cellulaire ou mérèse, qui a lieu dans les méristèmes.
L’augmentation des dimensions des cellules ou auxèse. Le plus souvent elle a une
direction :elle est soit longitudinale(élongation), soit radicale(croissance en épaisseur).
La différenciation des cellules. Les cellules indifférenciées sont des cellules capables de
donner n’importe quel type cellulaire végétal). Celles-ci prennent des caractères
morphologiques et physiologiques particuliers.
La mérèse a lieu dans les méristèmes. Il existe deux types de méristèmes. On trouve les
méristèmes primaires, situés à l’apex des racines, dans les bourgeons apicaux(à l’extrémité
des tiges et des rameaux) et axillaires(à l’aisselle des feuilles), et dans les entrenœuds
(méristèmes intercalaires). Les divisions se font de manière à aboutir à un massif plus ou
moins volumineux de cellules. Celles qui en dérivent, à la suite de leur allongement et de leur
différenciation, vont aboutir à des ébauches d’organes racinaires et de la tige, et assureront
leur croissance en longueur. On trouve également les méristèmes secondaires. Ils se
développent dans les organes les plus âgés. Ils permettent la croissance en épaisseur.
L’auxèse se traduit le plus souvent par une élongation des cellules ; Celle-ci est permise par la
composition des parois primaires et secondaires. La paroi primaire est essentiellement
constituée d’extensine(protéine de la paroi), d’hémicellulose, de composés pectiques et de
cellulose. La paroi secondaire donne une croissance par apposition, c’est-à-dire par dépôt de
couches successives.

4/23
Développement des organes végétatifs
Racines
La racine est, avec la tige et la feuille, l’une des composantes anatomique
fondamentales des végétaux. Celles-ci apparaissent comme l’organe le plus vital de la plante.
La racine est la prolongation de la partie basale de la tige. On peut observer une racine
principale, dite séminale. Celle-ci comporte une coiffe à son extrémité, une zone subterminale
où a lieu la croissance en longueur, et une zone de ramification dans laquelle apparaissent des
racines secondaires qui ont les mêmes caractéristiques que la racine principale.
Pour la croissance en longueur des racines on peut distinguer deux phases successives :la
multiplication des cellules puis l’élongation cellulaire.
La croissance en longueur de tout type de racine(principale ou secondaire) est assurée par un
méristème apical(situé à l’extrémité), localisé sous la coiffe. Les cellules produites dans cette
zone vont ensuite s’allonger dans la zone d’élongation, située quelques millimètres plus haut
que l’apex. Enfin, les cellules se différentient en tissus dans cette zone.
Bourgeons
Les bourgeons sont situés à l’apex des tiges ou des rameaux et à l’aisselle des feuilles.
Ils possèdent des jeunes feuilles imbriquées qui entourent le méristème apical par exemple.
Les bourgeons dit végétatifs assurent le développement des tiges et des rameaux végétatifs
avec leurs feuilles.
Tiges et Rameaux
La tige se développe à partir du bourgeon apical, les rameaux à partir des bourgeons
axilaires.
La zone d’élongation est repérable grâce à l’inégale longueur des entrenœuds. Sa longueur est
très variable selon les espèces. Les rameaux se développent à partir de bourgeons axilaires
situés à l’aisselle des feuilles.
Le développement de la tige et des rameaux se poursuit par l’organisation d’un méristème dit
caulinaire, et par des processus d’élongation et de différenciation des tissus.
Feuilles
La feuille présente une croissance en surface uniquement. Les zones qui produisent les
cellules et qui permettent leur accroissement sont moins délimitées que dans la tige et dans la
racine. Les feuilles ont une durée de vie limitée et finissent par tomber.
Le Cycle Cellulaire
Répétitif, il est constitué principalement de deux phases : l'interphase et la mitose.
Pendant l'interphase a lieu la réplication semi-conservative de l’ADN, tandis que la mitose est
la division cellulaire (répartition du matériel génétique dans deux cellules filles identiques).
Après un cycle cellulaire, on dit qu'il y a eu reproduction conforme de la cellule initiale. C'est
ce cycle cellulaire qui contribue au renouvellement moléculaire du matériel génétique. Il
comporte trois phases:
La phase G1 est la phase principale de la cellule. C'est pendant cette phase que se
déroulent les principales actions qui permettent de définir un type cellulaire et que la
cellule prend la morphologie caractéristique de son type. C'est durant cette période
que se déroule la croissance de la cellule.

5/23
La phase S, aussi appelée phase de synthèse, est la phase pendant laquelle la cellule
duplique son ADN. En fin de duplication, les deux chromosomes fils restent attachés
par une région appelée centromère. Chacun prend alors le nom de chromatide.
La phase G2 est intermédiaire entre la synthèse d'ADN et la mitose. Elle a été peu
étudiée, les mécanismes qui s'y déroulent sont mal connus.
La mitose
La mitose permet la répartition égale des chromosomes dans deux cellules filles. La
synthèse d'ADN à l'interphase ainsi que les modalités de répartition des chromosomes sont
semblables dans les cellules animales et végétales. La mitose se réalise en quatre étapes
successives : la prophase, la métaphase, l’anaphase et la télophase.
La Prophase
Lors de cette phase, le matériel génétique (ADN), qui en temps normal est présent
dans le noyau sous la forme de chromatides se condense en structure très ordonnée appelée
chromosome. Comme le matériel génétique a été dupliqué avant le début de la mitose, il y a
alors deux copies identiques de chaque chromosome dans la cellule. Ces chromosomes
identiques sont reliés l'un à l'autre par leur centromère. Lorsque les chromosomes sont ainsi
reliés, chaque chromosome individuel de la paire est appelé un chromatide, alors que l'unité
complète, de manière à entraîner une confusion, est nommée chromosome.
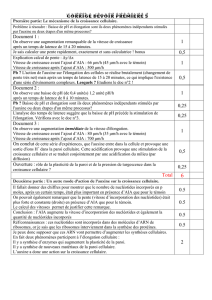
B – Le Rôle du Climat
a) Les effets de la Température
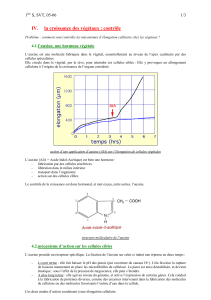
Nous avons observé, lors de nos expériences sur la croissance, que placer des plantes
dans un milieu à température basse ou élevée avait des conséquences sur le développement
des végétaux. A certaines températures elles poussaient relativement bien, tandis qu’à d’autres
leur croissance était inhibée. Nous avons pensées qu’il existait un intervalle de température
dans lequel la plante se développait normalement.
L’intervalle de température compatible avec une vie active est assez étroite :en général de –5
ou –10°C à +45°C. Toutefois ces limites sont largement reculées, car certains végétaux ont la
possibilité de passer à l’état de vie latente(activités cellulaires réduites au minimum). A
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
1
/
23
100%