INTRODUCTION A L`EMBRYOLOGIE

1
Module d’embryologie
I. INTRODUCTION A L’EMBRYOLOGIE
1. Définition
Le terme "embryologie" signifie l'étude de la formation et du développement des êtres vivants.
Toutefois, il est rare d'employer le terme dans le cas des êtres unicellulaires, même si ceux-ci peuvent
passer par différents stades durant leur vie. On parle de cycle cellulaire ou cycle vital. L'embryologie
suppose l'étude du développement d'êtres pluricellulaires, résultant de la reproduction sexuée de leurs
parents. Néanmoins, les eucaryotes multicellulaires ont évolué à partir de protozoaires unicellulaires et
c'est chez eux que sont apparus les caractères de base du développement.
Le sexe et la reproduction sont deux processus distincts et qui peuvent être séparés:
La reproduction consiste en la création de nouveaux individus;
Le sexe consiste en la combinaison de gènes de deux individus différents en un nouvel arrangement.
L'union des deux processus distincts, sexe et reproduction, en reproduction sexuée se rencontre pour la
première fois chez certains eucaryotes unicellulaires. Les individus sont morphologiquement identiques
(pas de mâles et de femelles) et haploïdes. Il s'agit d'une reproduction, car plusieurs nouveaux individus
sont formés.
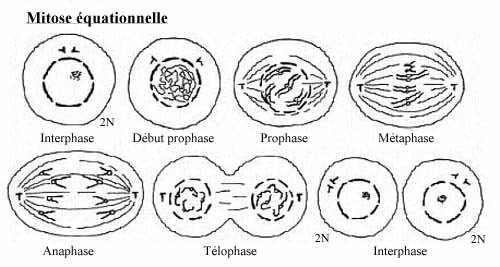
Les organismes pluricellulaires ont pour la plupart adopté la voie de la reproduction sexuée. Ils se
constituent à partir d'une cellule diploïde initiale, le zygote, par une succession de mitoses
équationnelles. Ce processus conduit à l'établissement d'une communauté formée, d'une part, d'un
grand nombre de cellules diploïdes qui sont spécialisées morphologiquement et fonctionnellement: la
lignée somatique, et, d'autre part, d'un autre type cellulaire diploïde qui évolue de façon particulière: la
lignée germinale. Au terme de leur évolution, les cellules de la lignée germinale entreprennent la
méiose et donnent naissance à des cellules haploïdes spécialisées en vue de la reproduction: les
gamètes. L'union des gamètes mâle et femelle, la fécondation, conduit à la formation du zygote
diploïde, qui est à l'origine d'un nouvel organisme.

2
2. Différents types d’œufs
On distingue plusieurs types d'œufs en fonction de leur structure (quantité de vitellus : jaune d’œuf):
Oligolécithe (oligos, peu nombreux) : Œuf qui contient une petite quantité de vitellus sous
forme de granulations dispersées dans le cytoplasme, il existe chez les échinodermes (les
oursins).
Hétérolécithe (heteros, autre) : Ce terme désigne les œufs dont le vitellus est réparti de façon
inégale, provoquant une segmentation inégale (Chez les amphibiens, certains plathelminthes, la
plupart des annélides et la plupart des mollusques).
Télolécithe (telos, fin) : Œuf caractérisé par un volume considérable de vitellus localisé à l’un
des pôles. La segmentation n'atteint que le territoire situé près du pôle animal (disque). Cette
segmentation produit une blastula en forme de disque (blastodisque), qui flotte sur les réserves
de vitellus. Il existe chez les oiseaux, les reptiles et la plupart des poissons.
Centrolécithe : Œuf dont le vitellus est surtout concentré au centre. Ce type d'œuf se rencontre
chez les insectes. Leur noyau est très légèrement excentré du fait de la présence de vitellus sous
forme de petites granulations dans le cytoplasme. Le noyau est au milieu de la quantité de
vitellus et le cytoplasme se divise en cytoplasme central et cytoplasme périphérique.
Alécithe (a, négation) : Œuf caractérisé par un volume très faible de vitellus (ou bien absence
total du vitellus). Les blastomères qui résultent de la segmentation (égale) seront sensiblement
égaux. Chez les mammifères placentaires, on donne aussi ce nom à un ovule non fécondé.
3. Segmentation (clivage)
L’ensemble de divisions mitotiques successives réalisées par le zygote pour former un corps cellulaire
appelé « Morula ». Ces cellules s’organisent dans une couche cellulaire « le blastula », suivi par des
mouvements cellulaires qui aboutissent à la mise en place des 3 feuillets embryonnaires «gastrula ».
La segmentation débute peu après la fécondation et se poursuit plus ou moins rapidement et plus ou
moins longtemps, selon les espèces.

3
La nature de l’ovule détermine le type de segmentation :
a. Segmentation totale (Holoblastique) : pour les ovules qui contiennent une petite quantité de
vitellus, le zygote se divise en 2, 4, 8, 16, 32, …..pour former le morula, cette segmentation peut être
égale : dans les ovules alécithes, les cellules résultantes (blastomères) ont la même taille, ou inégale :
chez les amphibiens (ovules hétérolécithes), les cellules (blastomères) du pole végétatif (PV) sont
plus volumineuses (chargées en vitellus) appelées : Macromères, et celles du pole animale (PA) sont
les moins volumineuses appelées : Micromères.
Les 2 premières divisions sont perpendiculaires, donnent 4 blastomères.
La 3ème division est horizontale et donne 8 blastomères : 4 au PA et 4 au PV.
La 4ème est perpendiculaire dans l’égale (8 au PA et 8 au PV), la 5ème est horizontale (16 au PA et 16 au
PV)…….. pour former enfin le morula puis le blastula. (Blastomères+blastocèle).
Dans l’inégale : après la 3ème division on obtient les 2 types de blastomères (micro et macro) dans le PA
et le PV pour former un morula et un blastula constitués des 2 types de blastomères.
b. Segmentation partielle (méroblastique) : on distingue 2 types de division :
* Discoïdale : dans les ovules télolécithes ; se déroule dans le disque qui renferme le noyau et le
cytoplasme.
* Superficielle : dans les ovules centrolécithes : la segmentation ne concerne que le noyau, il se divise
en 2, 4, 8…. un ensemble de noyaux, puis un grand nombre migre vers le cytoplasme périphérique pour
former une couche de cellules de blastomères, ce blastula se caractérise par l’absence de l’espace
interne (blastocèle).
4. Gastrulation :
L’ensembles des mouvements morphogénétiques réalisés par les cellules du blastula (les blastomères)
qui aboutissent à la formation des 3 feuillets (couches) embryonnaires : le feuillet externe : ectoderme,
le feuillet moyen : mésoderme et le feuillet interne c’est l’endoderme qui seront à l’origine de
l’organogenèse.

4
Il existe 4 types de gastrulations :
a. Gastrulation par invagination (Embolie) : dans les ovules oligolécithes, les cellules du PV
s’enfonce dans le blastocèle qui tente à réduire de volume puis à disparaître, il délimite une
cavité appelé l’archenteron qui est à l’origine de la formation de l’intestin, qui ouvre à
l’extérieure par la blastopore, c’est la mise en place du tube digestif. Enfin l’endoderme limite
l’archenteron, le mésoderme borde cette cavité et l’ectoderme la couche externe.
b. Gastrulation par recouvrement (Epibolie) : dans les ovules hétérolécithes, les cellules du PV
sont trop volumineuses pour d’enfoncer dans le blastocèle, donc de manière passive les cellules
du PA se multiplient pour recouvrir les cellules du PV pour former l’ectoderme, et les cellules
du PV forment l’endoderme.
c. Gastrulation par immigration : chez les vertébrés supérieurs, ex : les oiseaux, les cellules du
blastodisque migrent dans le blastocèle pour former l’endoderme puis le mésoderme.
d. Gastrulation par délamination : dans les ovules centrolécithes, les cellules du blastomères se
divisent pour former une 2ème couche (endoderme) et la couche externe donne l’ectoderme.
5. Neurulation : la neurulation marque le début de l'organogenèse.
Chez les Vertébrés, la gastrulation a créé un embryon ayant un feuillet ectodermique externe, un
feuillet mésodermique intermédiaire et un feuillet endodermique interne. En plus, un cordon de
cellules, la notochorde, se trouve juste sous la portion dorsale de l'ectoderme. La notochorde exerce un
effet inducteur sur l'ectoderme sous-jacent pour qu'il forme le tube neural: il s'agit de l'induction
embryonnaire primaire.
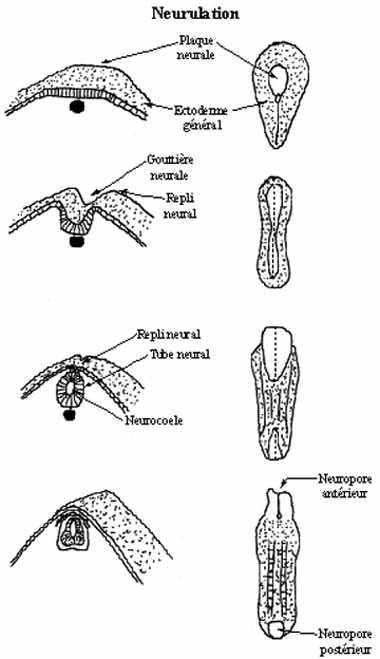
L'embryon sur le point de neuruler s'allonge dans l'axe antéro-postérieur et s'aplatit dorsalement. Le
premier indice qu'une région de l'ectoderme devient spécifiée à devenir tissu neural se manifeste par un
changement de la forme des cellules de l'ectoderme dorsal de l'embryon. Ces cellules s'allongent,
devenant cylindriques, tandis que les autres cellules ectodermiques sont cuboïdales ou pavimenteuses.
Les cellules cylindriques dorsales forment ensemble le neurectoderme: épithélium simple (une seule
assise de cellules) cylindrique (les cellules qui le composent sont toutes cylindriques). Le
neurectoderme étant surélevé par rapport au reste de l'ectoderme (ectoderme général), on l'appelle à ce
stade plaque neurale.

5
Près de 50% des cellules ectodermiques sont inclues dans la plaque neurale.
Les rebords de la plaque neurale, à la limite neurectoderme - ectoderme général, s'épaississent et se
soulèvent: les replis neuraux, entre lesquels se crée une dépression en forme de U: la gouttière
neurale.
Le tube neural est à l'origine du système nerveux central (cerveau et moelle épinière), et la crête
neurale de presque tout le système nerveux périphérique et d'une bonne partie du squelette crânio-
facial.
La neurulation ne se produit pas simultanément sur toute la longueur de l'embryon. Amorcée à la
jonction tête - tronc, elle se poursuit antérieurement et postérieurement. Chez le poulet de 24 heures
d'incubation, elle est fort avancée dans la région de la tête tandis que la région caudale est encore en
gastrulation. Chez les amphibiens la neurulation s'effectue en 24 heures à 18°C. Chez les oiseaux elle
débute vers la 20e heure d'incubation. L'embryon des Vertébrés ayant subi ces changements s'appelle
neurula.
1
/
5
100%












{kind=link}
{kind=link}