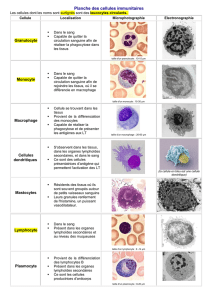
II. Les érythrocytes

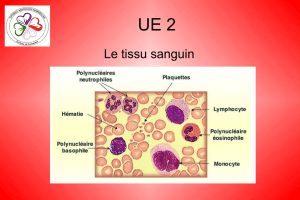
Tissu sanguin
Tissu sanguin ................................................................................... 1
I. Généralité ............................................................................... 2
A. Aspect quantitatif ................................................................ 2
1) Généralité ....................................................................... 2
2) Formule leucocytaire ........................................................ 2
B. Aspect qualitatif .................................................................. 3
C. Les différentes lignés cellulaire ............................................. 3
1) Origine des cellules sanguines ........................................... 3
2) Technique ayant permis la découverte de l’origine des cellules
sanguines ................................................................................ 3
3) Cellules souches totipotentes ............................................. 3
4) Cellules multipotentes ....................................................... 4
II. Les érythrocytes .................................................................... 6
A. Erythropoïèse ..................................................................... 7
1) De la cellule progénitrice myéloïde au proérythroblaste ......... 7
2) Du proérythroblaste à l’érythrocyte .................................... 7
(a) Evolution ...................................................................... 7
(b) Aspect des différentes lignées ......................................... 7
3) Contrôle de l’érythropoïèse ................................................ 9
B. Morphologie et caractéristiques des érythrocytes .................... 9
1) Au microscope optique ...................................................... 9
2) Au microscope électronique ............................................... 9
3) Aspect quantitatif ............................................................. 9
4) Membrane du globule rouge ............................................ 10
C. Anomalie de l’érythrocyte .................................................. 10
III. Les thrombocytes ................................................................ 11
A. Morphologie ..................................................................... 11
1) Au microscope optique .................................................... 11
2) Au microscope électronique ............................................. 11
3) Aspect qualitatif ............................................................. 12
B. Rôle ................................................................................ 12
C. Maturation des thrombocytes ............................................. 13
IV. Les polynucléaires neutrophiles ............................................. 15
A. Morphologie ..................................................................... 15
B. Genèse du polynucléaire .................................................... 16
C. Rôles ............................................................................... 16
V. Polynucléaire éosinophile ...................................................... 17
A. Morphologie ..................................................................... 18
B. Genèse ............................................................................ 18
C. Rôles ............................................................................... 19
VI. Polynucléaire basophile ........................................................ 19
A. Morphologie ..................................................................... 19
B. Rôles ............................................................................... 20
VII. Mastocytes ......................................................................... 20
A. Morphologie ..................................................................... 20

VIII. Monocytes ....................................................................... 21
A. Rôles ............................................................................... 21
IX. Lymphocytes ....................................................................... 22
A. Généralité ........................................................................ 22
B. Les lymphocytes B ............................................................ 22
C. Les lymphocyte T .............................................................. 23
D. Lymphopoïèse .................................................................. 23
X. Les organes lymphoïdes ....................................................... 24
A. La moelle osseuse ............................................................. 24
B. Le ganglion lymphatique .................................................... 24
C. La rate ............................................................................ 28
D. Le thymus ........................................................................ 30
E. Les formations lymphoïdes .................................................... 32
1) Les amygdales ............................................................... 33
2) L’appendice ................................................................... 33
3) Plaque de Payer ............................................................. 33
I. Généralité
Le sang est constitué de cellule en suspension dans du plasma. Les
cellules sanguines représentent 40% du volume total, c'est l’hématocrite.
La formule sanguine détermine la proportion des différentes variétés de
globule blanc.
Pour étudier, on fait un frottie sanguin coloré au MGG qui permet de
distinguer les différentes variétés de globule blanc.
L’hémogramme varie en fonction de l’age.
A. Aspect quantitatif
1) Généralité
Il y a environ 4 à 5 millions de globule rouge par ml. La concentration en
globule rouge est supérieure chez les hommes que chez les femmes. Dans
la circulation sanguine, il y a des réticulocyte qui sont des globule rouge
immature et représentent 0,5%, c'est-à-dire 25000 cellules par ml.
Les globule blanc ou leucocytes représentent 4000 à 10000 cellules par
ml.
Les plaquettes représentent 150 à 400000 cellules par ml.
2) Formule leucocytaire
La formule leucocytaire va être exprimé soit en % ou en nombre de
cellules.
Les granulocyte ou polynucléaire vont être distinguer en trois variété par
le MGG qui mettra en évidence les granulations. Ces granulations peuvent
être neutrophiles, ce type représente 2000 à 7500 cellules. Elles peuvent

être aussi basophile représentant 0 à 200 cellules et éosinophile
regroupant 40 à 700 cellules.
Les lymphocytes représentent 1000 à 4000 cellules.
Les monocytes représentent de 200 à 1000 cellules.
B. Aspect qualitatif
La taille des globule rouge est d’environ 7 µm. Les lymphocyte ont une
taille de 12µm. Les polynucléaires seront plus volumineux avec leur aspect
particulier du noyaux. Les cellules les plus grandes sont les monocytes.
C. Les différentes lignés cellulaire
1) Origine des cellules sanguines
L’origine des cellules sanguines est la moelle osseuse où vont se situer
les cellules souches. La production des ces cellules sanguines se fait au
cours de l’hématopoïèse à partir de la moelle hématogène. Cette moelle
hématogène comporte trois compartiments. On a des cellules souches qui
sont dites totipotentes. Elles ont deux propriétés, elles sont capables
d’autorenouvelement et elles vont donner également des cellules qui
entrent en différenciation. Le deuxièmement compartiment va être
constituer par des cellules souches multipotentes ou des progéniteur. Le
troisième compartiment, c'est celui des cellules déterminées, c'est-à-dire
des cellules qui vont entrer en différenciation terminale. Ces précurseurs
ne sont pas capables de autorenouvelement.
2) Technique ayant permis la découverte de
l’origine des cellules sanguines
Cette cellule souche a été découverte en 1961 sur la souris. Les
rongeurs ont une hématopoïèse médullaire et splénique. Ils ont irradiés
des souris ce qui entraînent une aplasie médullaire, c'est-à-dire une
disparition des cellules hématopoiétique. Si a ce moment on injecte à
l’animal des cellules de moelles osseuse d’une autre sourie syngénique
apparaît au sein de la rate des colonie cellulaires qui étaient des colonies
mixtes avec les différentes cellules des différentes lignés. On a appelé ces
cellules formant ces colonies les CFU - S.
Une autre manière de le montrer est l’étude de certaine pathologie. Chez
les patient qui présent un déficit des enzymes qui existent sous deux
isoforme exprimé chez les femme. Si il y a une pathologie sanguine les
cellules anormales exprimeront toutes le même isoformes, il y a donc une
cellule dont dérive toutes les autres.
3) Cellules souches totipotentes
Ces cellules souche totipotente n’ont pas de caractéristique particulière
et ressemble beaucoup à des lymphocytes. On peut les identifier par des
clusters de différenciation qui varie en fonction de l’évolution de la cellule.
On sait que la cellule souche totipotente exprime le CD34 qui

progressivement va être associé à d’autre marqueur et premier la CD34
lorsque elles devient
totipotente.
Représentation schématique des différents compartiments du système hématopoïétique
Moelle
osseuse
Cellules souches Cellule dendritique
Pro-Monocyte Monocyte
Progéniteur
éosinophile
Progéniteur
neutrophile
Progéniteur
basophile
neutrophile
éosinophile
basophile Mastocyte
Macrophage
BFU-MK Mégacaryocyte Plaquettes
Érythroblaste Érythrocyte
Pro-NK
Pro-T
NK
Cellule dendritique
Pré-T T auxiliaire
T cytotoxique
sang
Pro-B Pré-B Lymphocyte B Plasmocyte
CLP
CMP
EMP
GMP
BFU-E
CSH Progéniteurs Cellules matures
4) Cellules multipotentes
Les cellules multipotentes sont au nombre de deux. Il y a les
progéniteurs myéloïdes et les progénitures lymphoïdes.
Les progéniteurs lymphoïdes vont être à l’origine des lymphocytes et les
progéniteurs myéloïdes vont être à l’origine de toute l’autre cellule.
(Globule rouge, polynucléaire, monocytes, plaquettes).

 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
1
/
34
100%