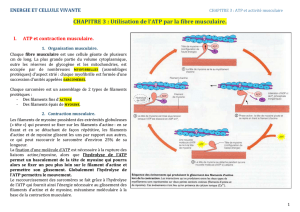
les constituants de la chaine respiratoire

1
Bioénergétique
Introduction :
La bioénergétique ou thermodynamique biochimique est l’étude des variations d’énergie
associées aux réactions biochimiques. Elle fournit les principes fondamentaux expliquant
pourquoi certaines réactions peuvent se produire alors que d’autres pas. Les systèmes non
biologiques peuvent utiliser l’énergie calorifique pour effectuer un travail, mais les systèmes
biologiques sont essentiellement isothermes et utilisent l’énergie chimique pour faire
fonctionner les processus vitaux.
Energie libre
Dans un système, la variation de l’énergie libre selon Gibbs (ΔG) est la portion de la variation
de l’énergie totale disponible pour accomplir le travail, elle correspond à l’énergie utile
connue dans les systèmes chimiques comme étant le potentiel chimique.
Les systèmes biologiques sont conformes aux lois générales de la thermodynamique
Le premier principe de la thermodynamique établit que l’énergie totale d’un système et de son milieu
environnant demeure constante.
C’est aussi la loi de la conservation de l’énergie. Dans un système total, l’énergie n’est ni perdue ni
gagnée durant une transformation. Dans ce système total, l’énergie pourrait être transférée d’une
partie à une autre ou transformée en une autre forme d’énergie.
Exemple : chez les organismes vivants, l’énergie chimique peut être transformée en chaleur, en
énergie électrique, en énergie radiante ou en énergie mécanique.
Soit une réaction chimique :
A + B C + D (réaction 1).
Les molécules A, B ,C et D ont une énergie libre ( énergie de vibration ,rotation et translation de la
molécule , énergie des liaisons interatomiques ) . l’énergie libre est appelée GA , GB , GC et GD.
La différence d’énergie libre entre C et D (état final) d’une part et A et B (état initial) d’autre part est
appelée variation d’énergie libre ΔG :
ΔG = (GC + GD)-(GA + GB) (état final – état initial)
ΔG est exprimé en kilojoules/ mole (KJ.mol-1) , ou anciennement en Kilocalories/mole. (Kcal.mol-1).
Le second principe de la thermodynamique stipule que l’entropie totale d’un système doit augmenter
si un processus s’effectue spontanément.

2
L’entropie représente le niveau de désordre ou de chaos d’un système et devient maximum dans un
système qui s’approche de l’équilibre vrai.
Dans des conditions de température et de pression constantes, la relation entre la variation d’énergie
libre (ΔG) d’un système en réaction et la variation de l’entropie (ΔS) est donnée par l’équation
suivante qui combine les deux lois de la thermodynamique (équation de Gibbs) :
ΔG = ΔH- TΔS
Où ΔH est la variation de l’enthalpie (chaleur), et T la température absolue.
Dans les conditions des réactions biochimiques, ΔH est à peu près égal à ΔE (la variation totale de
l’énergie interne de la réaction).
La relation citée peut s’exprimer de la façon suivante :
ΔG = ΔE –TΔS
Si ΔG est négatif, la réaction procède spontanément avec perte d’énergie libre, c'est-à-dire qu’elle est
exergonique.
Si ΔG est élevé, la réaction se fait pratiquement jusqu’à la fin et elle est essentiellement irréversible.
Si ΔG est positif, la réaction ne se produit que s’il y a gain d’énergie libre, elle est dite réaction
endergonique.
Si ΔG est élevé, le système est stable et la réaction n’a pas tendance à se produire.
Si ΔG est égal à 0, le système est à l’équilibre et aucune variation nette ne se produit.
La variation standard de l’énergie libre
Quand les réactifs sont présents à des concentrations de 1.0 mol/l, ΔG0 représente la
variation standard de l’énergie libre.
Pour les réactions biochimiques, l’état standard est défini comme ayant un pH de 7.0.
La variation standard de l’énergie libre dans ces conditions est appelée ΔG0’.
La variation standard de l’énergie libre peut être calculée à partir de la constante d’équilibre
K’eq.
ΔG0’ = -2.303 RTlog K’eq
Où R est la constante des gaz parfaits, T la température absolue.
K’ eq rapport des constantes de vitesse, c’est la constante d’équilibre d’une réaction, dans la
réaction1 K’eq = (C )eq ( D )eq /(A )eq (B)eq
Remarque : la véritable ΔG pourrait être plus grande ou plus petite que ΔG0’ selon les
concentrations des divers réactifs, solvant, ions et les protéines.

3
Dans les conditions réelles de la vie cellulaire, la température est proche de 37°C, les
concentrations initiales des produits et réactifs sont très inferieures à 1M .On démontre que :
ΔG = ΔG0’ + RTln (C) (D) /(A) (B),
où (C) (D) (A) (B) sont les concentrations initiales des réactants. ΔG n’est donc pas une
constante.
5 conséquences pratiques sont tirées à partir de la loi démontrée (citée plus haut) pour le
calcul de ΔG :
- au fur est à mesure que la réaction A +B C+D évolue, RTln (C) (D) /(A) (B)
augmente, la valeur absolue de ΔG s’annule lorsque l’équilibre est atteint.
ΔG0’ = -RTlnKeq. La constante d’équilibre d’une réaction permet de calculer sa variation
d’énergie libre standard ΔG0’.
Plus la valeur d’une ΔG0’négative est grande, plus l’équilibre s’établit loin du côté des
produits, plus la réaction est complète dans les conditions standard.
Exemple : soit la réaction : ATP +H2O ADP +Pi ΔG0’ de la réaction d’hydrolyse
de l’ATP est de -30.5 KJ.mol-1.
- connaissant la ΔG0’et les concentrations initiales des réactifs et produits dans des conditions
cellulaires données, on peut calculer ΔG :
ΔG = ΔG0’ +RTln( C) ( D )/( A) ( B )
Exemple la ΔG de la réaction d’hydrolyse de l’ATP dans la cellule est de ; -50KJ/mol
La réaction d’hydrolyse de l’ATP est donc plus exergonique dans les conditions cellulaires
que dans les conditions standard.
- une réaction dont la ΔG0’ est positive (donc impossible dans les conditions standard) peut
avoir lieu pour certaines concentrations des réactants, quand ( C) ( D ) /( A) ( B) est inférieur
à 1 , la ΔG est négatif et la réaction est possible.
Exemple, la réaction d’oxydation du lactate en pyruvate, a une ΔG0’de 27 KJ/ml.
Lactate + NAD+ pyruvate + NADH,H+
-une réaction est d’autant plus réversible, donc peut fonctionner dans un sens ou dans l’autre
(par une modification des concentrations des réactants) que sa ΔG0’ est plus proche de zéro.
-une réaction endergonique ( ΔG0’ positive) est possible si elle est couplée à une réaction
exergonique ( ΔG0’ négative).
Exemple, la réaction : glucose +phosphate glucose-6-phosphate a une ΔG0’
de +13.8KJ/mol.

4
La réaction est thermodynamiquement défavorable dans les conditions standard .pour qu’elle
puisse se produire de l’énergie est fournie par couplage à une autre réaction qui elle est
exergonique, c’est la réaction d’hydrolyse de l’ATP en ADP et Pi dont la ΔG0’ = -30.5KJ/mol.
La réaction global est exergonique, sa ΔG0’ = -16.7KJ/mol.
L’enzyme qui catalyse la double réaction est hexokinase, glucokinase.
Remarque : dans un système de réactions biochimiques, une enzyme ne fait que rendre
l’équilibre plus rapidement accessible.
Energie libre d’activation ΔG#
La vitesse d’une réaction dépend de l’énergie libre d’activation ΔG# qui est différente de ΔG
(ΔG ne donne aucune information sur la vitesse de réaction, elle informe dans quel sens a lieu
spontanément une réaction, combien de travail elle fournit).
Soit la réaction : A +B C + D
L’état initial A + B à haute énergie libre, est séparé de l’état final C +D à basse énergie
libre, par un état de transition d’énergie libre supérieure à celle de l’état initial, la différence
d’énergie libre entre l’état initial et l’état de transition est l’énergie libre d’activation ΔG#.
La vitesse de la réaction est inversement proportionnelle à ΔG#.
Dans la cellule ce sont les enzymes qui accélèrent la réaction, ils abaissent l’énergie libre
d’activation .
Les réactions endergoniques procèdent par couplage à des réactions exergoniques
Les réactions de synthèse, la contraction musculaire, la conduction de l’influx nerveux et le
transport actif, obtiennent l’énergie par liaison chimique ou couplage aux réactions
d’oxydation.
A + C B + D + chaleur
La conversion du métabolite A en métabolite B s’effectue avec une libération d’énergie libre.
Cette conversion est couplée à une autre réaction, dans laquelle l’énergie libre est requise pour
transformer le métabolite C en métabolite D. Etant donné qu’une partie de l’énergie libérée
dans la réaction de dégradation est transférée à la réaction de synthèse sous une autre forme
que la chaleur, on dit : un processus exergonique et endergonique.
En pratique, un processus endergonique ne peut exister indépendamment, mais doit faire
partie d’un système couplé exergonique-endergonique.
Les réactions exergoniques sont groupées sous le terme de catabolisme (dégradation et
oxydation des molécules combustibles), et les réactions de synthèse des molécules, sous le
terme d’anabolisme.

5
Un mécanisme possible de couplage pourrait être envisagé si un intermédiaire obligatoire
commun participait aux deux réactions :
A + C I B + C
Il faut noter que ce type de couplage possède un mécanisme intrinsèque de contrôle
biologique (contrôle respiratoire), qui empêche un organisme de bruler de façon incontrôlée.
Une extension de ce concept de couplage est fourni par les réactions de déshydrogénation qui
sont couplées à des hydrogénations par un transporteur intermédiaire.
Une autre alternative de couplage d’une réaction exergonique à une réaction endergonique,
passe par la synthèse d’une molécule d’énergie potentielle élevée dans la réaction
exergonique et l’incorporation de cette substance nouvelle dans la réaction endergonique
(transfert d’énergie libre).
Dans la cellule, le principal intermédiaire ou transporteur hautement énergétique appelé ~ €
est l’adénosine triphosphate (ATP).
Résumé
Les activités des organismes vivants exigent de l’énergie. Les mouvements, la croissance, la
synthèse des biomolécules et le transport des ions à travers la membrane demandent un apport
énergétique. Tous les organismes doivent obtenir de leur environnement l’énergie dont ils ont
besoin. L’étude de ces phénomènes bioénergétique exige une connaissance de la
thermodynamique , un ensemble de principes décrivant les flux et les échanges de chaleur ,
d’énergie et de matière dans les systèmes considérés ainsi que la connaissance des principes
de base de la thermodynamique ; l’analyse des flux thermiques , la production de l’entropie ,
le rôle de l’énergie libre , quelques concepts annexes tels que ; état standard, effet du pH sur
l’énergie libre standard , effet de la concentration sur la variation nette de l’énergie libre d’une
réaction , importance des réactions couplées dans les processus métaboliques.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
1
/
37
100%