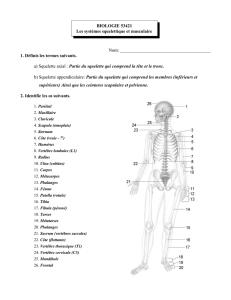
Physiologie de la contraction musculaire: Muscles squelettiques

Kevin CHEVALIER 1
PHYSIOLOGIE DE LA CONTRACTION MUSCULAIRE
I. Les muscles squelettiques
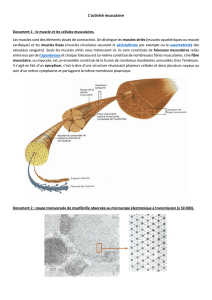
A. Structure du muscle
Le muscle est entouré de membranes conjonctives qui
délimitent des compartiments :
La plus externe est l'épimysium qui va se poursuivre de part et
d'autre par les tendons. Ces tendons se fixent sur le périoste
osseux.
Le périmysium divise le muscle en faisceaux et isole les fibres
musculaires des autres fibres musculaires.
Le périmysium donne naissance à l'endomysium constitué de
fines cloisons de tissu conjonctif lâche qui pénètrent dans les
faisceaux et entourent individuellement chaque fibre
musculaire.
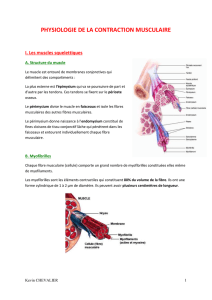
B. Myofibrilles
Chaque fibre musculaire (cellule) comporte un grand nombre de myofibrilles constituées elles même
de myofilaments.
Les myofibrilles sont les éléments contractiles qui constituent 80% du volume de la fibre. Ils ont une
forme cylindrique de 1 à 2 µm de diamètre. Ils peuvent avoir plusieurs centimètres de longueur.

Kevin CHEVALIER 2
C. La cellule musculaire (myocyte)
1. Ultrastructure de la cellule
Ce sont des cellules qui sont constituées par la fusion de plusieurs cellules jeunes (les myoblastes).
Ce sont donc des cellules polynucléées (formant donc un syncytium).
Les myocytes (ou myotubes) contiennent tant de myofibrilles que les noyaux sont repoussés à
l'extérieur.
Une myofibrille est formée de la succession d'éléments fondamentaux appelés sarcomères.
2. Triade
a. Tubules transverses
Les tubules transverses sont des invaginations de la membrane plasmique qui s'enfonce en
profondeur (en jaune).
L'ensemble des tubules transverses constitue, par définition, le système T. Il y a 2 tubules
transverses par sarcomère.

Kevin CHEVALIER 3
b. Réticulum sarcoplasmique
Le réticulum sarcoplasmique (lisse) est très développé dans le muscle squelettique. Il constitue des
citernes, parallèle aux myofibrilles. Ils s'accolent aux tubules transverses. C'est un réservoir de Ca2+.
Deux citernes terminales s'associent avec un tubule T pour former une triade.
D. Sarcomère
Un sarcomère est constitué : de 2 demi-disques I et d'un disque A. Il se situe donc entre deux
bandes Z.
Le disque I (clair) est hétérogène. Il est divisé en deux par la bande Z.
Le disque A (sombre) est hétérogène également et possède en son milieu une bande plus claire : la
bande H (ou AH) au milieu de laquelle se trouve la ligne M.

Kevin CHEVALIER 4
1. Filaments et cellule
Chaque myofibrille est constituée de myofilaments épais et fins.
La bande A est constituée d'un empilement de myofilaments épais.
Les myofilaments fins forment le disque I.
Les filaments fins s'ancrent au milieu du disque I, formant la ligne Z. Ainsi la ligne Z est le point
d'ancrage des filaments fins les uns par rapport aux autres.
Ces myofilaments fins ont la propriété de glisser entre les filaments épais.
Dans un disque A on observe une superposition des filaments épais et des filaments fins dans la zone
AI.
Par contre la partie centrale n'est formée que de myofilaments épais. C'est la partie AH.
2. Contraction
Quand un muscle va se contracter, le sarcomère va se raccourcir. Les filaments eux ne changent pas
de taille : ils vont glisser les un sur les autres.
Durant la contraction, les myofilaments fins glissent entre les filaments épais. Ainsi, la bande AH va
diminuer. Et comme une myofibrille est constituée d'une multitude de sarcomère, elle va se
raccourcir, la cellule va se raccourcir et donc le muscle va se raccourcir si la contraction est
isotonique.

Kevin CHEVALIER 5
3. Etape moléculaire
a. Filaments fins
Ce sont des filaments d'actine qui sont formés d'une multitude de molécule d'actine G.
L'actine G (globulaire) s'associe en polymère fibreux qui forme l'actine F (fibrillaire) sous la forme de
deux filaments qui s'enroulent l'un autour de l'autre.
On a aussi la présence de tropomysine qui est localisée dans le sillon central entre les 2 filaments
d'actine.
Il y a aussi présence de troponine qui est sous trois formes :
La troponine C qui fixent le calcium
La troponine T qui se lie à la tropomyosine. Ainsi elle fixe la troponine à la tropomyosine.
La troponine I qui inhibe les interactions actine-myosine
b. Filaments épais
Ils sont constitués essentiellement par des molécules de myosine
C'est une protéine constituée de 6 polypeptides avec 2 chaînes lourdes
(avec une partie globulaire) et de 4 chaînes légères qui sont attachées
aux chaînes lourde au niveau de la partie globulaire.
Ainsi quand on parle d'une molécule de myosine, on va distinguer 3 parties :
La queue
Le cou
La tête
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%