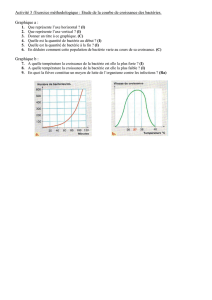
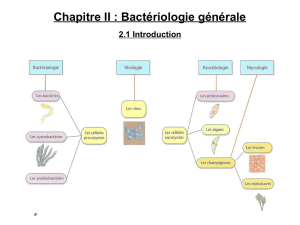
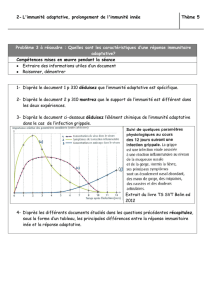
On commence ce cours par des géneralités : les différentes étapes

RT : LAROZE Amandine
Cours d’immuno du 18 mars 2008 de 15h à 17h
Toutes les diapos concernant ce cours sont sur intranet
DEFENSES ANTI-BACTERIENNES
On commence ce cours par des généralités : les différentes étapes du processus infectieux
La pathogénie des agents infectieux
Les facteurs de défense de l’hôte
Puis ensuite on verra les deux grands types de défense immunitaire qui interviennent dans la
défense antibactérienne, et plus particulièrement l’immunité innée non spécifique avec la mise
en jeu des barrières anatomiques et les réactions inflammatoires. Ensuite la réponse adaptative
de l’hôte contre l’infection et on verra à la fin quels sont les acteurs qui interviennent contre
les bactéries intracellulaires et extracellulaires.
I. généralités :
Etapes du processus infectieux:
Pour qu’une bactérie pénètre dans l’organisme, il faut qu’elle franchisse la barrière épithéliale
cela est possible seulement s’il y a une brèche cutanée ou muqueuse (la bactérie ne franchit
pas la peau intacte).
Ensuite on a une extension du processus infectieux soit par voie directe soit de cellule en
cellule.
On aura une guérison spontanée ou sous traitement avec une élimination de l’agent bactérien
avec éventuellement la survenue de séquelles, de lésions tissulaires comme en cas de
survenue de granulome dans le cas de la tuberculose. Et éventuellement la possibilité de
réinfection.
Caractéristiques des agents pathogènes :
Les bactéries se caractérisent par :
Mode de transmission
Mécanisme de réplication
Pathogénie
Réponse immune induite, qui varie en fonction du type d’agent pathogène et du
type de bactérie impliquée.
Mécanisme de réplication des agents infectieux :
En ce qui concerne les bactéries extracellulaires elles ont la caractéristique de se répliquer en
dehors de la cellule, elles sont assez souvent enveloppés d’une capsule polysaccharidique ce
qui fait qu’elles vont résister à la phagocytose par les macrophages et les polynucléaires

neutrophiles donc il y aura mis en jeu d’autre éléments de la réponse immunitaire et en
particulier une opsonisation par les anticorps qui est nécessaire à leur dégradation.
Pour les bactéries intracellulaires, elles se répliquent soit dans la cellule soit dans les vésicules
cytoplasmiques.
Facteurs de virulence des bactéries extracellulaires :
La présence de molécules d’adhérences (permet d’adhérer aux différents épithéliums)
La présence d’une capsule
La production éventuelle de toxines
Leur variation antigénique qui pose le problème de réinfection
La présence de protéines de surface qui sont la cible des anticorps mais qui peuvent
également inhiber les fonctions des cellules phagocytaires.
Résistance des bactéries à l’environnement extracellulaire :
Certaines bactéries présentent la capacité de résister à l’environnement extracellulaire. En
particulier celles qui possèdent une capsule et des pili qui ont la capacité d’inhiber les
fonctions des cellules phagocytaires. Et également celles qui produisent des enzymes qui vont
entraîner une lyse des phagocytes ou une altération du chimiotactisme en particulier par la
C5a peptidase, car on verra que le C5a, qui est un dérivé du complément, a un fort pouvoir
chimiotactique c'est-à-dire qu’il a tendance à attirer vers le site de l’infection les cellules
phagocytaires.
Pathogénie des agents infectieux :
Ces différentes bactéries peuvent exercer leur pouvoir pathogène par différents mécanismes :
Soit par un mécanisme direct :
-par la production de toxines : exotoxines ou endotoxines (les anticorps luttent
contre cette production de toxines)
-ou par un effet cytopathique direct.
Soit par un mécanisme indirect : dans ce cas là les bactéries n’agissent pas par production de
ces toxines mais leur pathogenicité intervient par la mise en jeu d’un certain nombre de
réactions immunitaires comme la formation de complexes immuns, la formation d’auto-
anticorps ou le développement d’une immunité à médiation cellulaire.
Immunité anti-infectieuse : deux stratégies de défense.
L’introduction d’une bactérie dans l’organisme déclenche 2 types de réponses immunitaires :
-une réponse immunitaire inné ou naturelle.
-une réponse adaptative.
1. immunité innée :

commune à l’ensemble des organismes pluricellulaires
mise en jeu immédiate : défense précoce
ne reconnaît que des substances microbiennes et en particulier elle
reconnaît des PAMPs (Pathogen Associated Molecular Patterns) qui sont
des motifs communs à différentes familles de microorganismes. Cette
reconnaissance va entraîner un signal de danger qui va lui-même engendrer
la mise en jeu de la réaction inflammatoire.
Stimulation de l’immunité adaptative.
2. immunité adaptative : spécifique, acquise
se développe plus lentement, au bout de quelques jours.
défense tardive
réponse amplifiée et soutenue
induit une mémoire immunitaire donc les réponses ultérieures seront
déclenchées plus rapidement et seront moins importantes.
Met en jeu LB et LT portant des récepteurs d’antigènes spécifiques
Peut reconnaître des antigènes non microbiens
Il y a bien sur des coopérations entre les deux systèmes.
Deux types de réponse immunitaire :
Cette diapo est là pour nous montrer les différents acteurs qui interviennent dans les deux
types de réponses immunitaires.
La réponse immunitaire innée met en jeu en premier lieu les barrières anatomiques de
l’organisme, puis des facteurs solubles comme les interférons et le système du complément.
Et enfin il peut y avoir intervention de cellules comme les phagocytes (en particulier
les polynucléaires neutrophiles) mais également des cellules cytotoxiques comme les cellules
NK, et les cellules dendritiques qui font en fait le lien entre la réponse immunitaire innée et
adaptative.
La réponse immunitaire adaptative qui met donc plusieurs jours à s’installer et qui confère
une mémoire immunitaire. Elle fait intervenir les lymphocytes T, les lymphocytes B et bien
sur la production d’anticorps.
II. Immunité innée non-spécifique :
1. les barrières anatomiques :
Les barrières anatomiques font intervenir les barrières épithéliales puisqu’en fait les bactéries
ne franchissent pas la peau intacte. Il y a différents mécanismes qui permettent à ces barrières
anatomiques d’intervenir :
actions mécaniques : les bactéries sont exclues par desquamation de la couche
de keratinocytes et par le faible degré hygrométrique de cette couche. Il y a

également des phénomènes statiques par des jonctions très étroites entre les
cellules.
La sécrétion de mucus qui va inhiber l’adhérence des microorganismes ;
Des phénomènes dynamiques comme le péristaltisme ou encore la présence de
cils qui protégent la muqueuse des bactéries
actions chimiques : avec la présence d’acides gras qui sont toxiques pour la
bactérie, mais également la production d’enzymes comme le lysozyme, les
pepsines ou encore les défensines. Et enfin un certain nombre de bactéries sont
détruites par le pH acide notamment au niveau de l’estomac et au niveau du
vagin.
Actions microbiologiques : par la flore commensale qui va produire des
protéines antibactériennes.
Intervention d’IgA sécrétoires : permettent de neutraliser l’adhérence des
bactéries au niveau de l’épithélium, qui ont également une action opsonisante
et qui active la voie alterne du complément.
Parmi ces différents peptides antimicrobiens qui interviennent au niveau de ces barrières
anatomiques, il faut quand même situer le rôle des défensines :
-qui ont des effets antibactériens extrêmement importants
-qui sont produites par les cellules épithéliales :
→ les α défensines produites par les cellules de Paneth
→ les β défensines produites par les cellules épithéliales de la
muqueuse buccale, trachéale et des bronches.
Il y a également d’autres peptides antimicrobiens qui interviennent et certains sont produits
par le polynucléaire neutrophile, on y reviendra tout à l’heure.
Rôle des défensines : ce rôle a été démontré en cas d’infection à Staphylococcus
aureus, celui-ci produit de la staphylokinase qui explique sa résistance aux défensines
humaines. Ce rôle a été également évoqué dans la maladie de Crohn, qui est
caractérisé par la présence de mutations de la protéine NOD2 mais également par la
diminution de l’expression des α défensines.
Ces barrières épithéliales peuvent être mis en défaut en cas de présence de pores, de
follicules pileux et de glandes sébacées car dans ce cas là les conditions ne sont plus
optimales, mais également an cas de plaies, de brûlures, de présence de cathéters ou de
macération.
Une fois que les bactéries ont franchi ces différentes barrières, elles vont déclencher la
réponse immunitaire innée (qui se développe extrêmement rapidement), qui font intervenir
des facteurs solubles et des cellules phagocytaires : le polynucléaire neutrophile et le
monocyte.

La réponse va avoir lieu très rapidement puisque en une quinzaine de minutes la
bactérie par la voie alterne du complément va induire la production de C3a et de C5a. Ces
dérivés du complément vont venir agir sur la barrière endothéliale en augmentant la
perméabilité vasculaire ce qui va faciliter la migration des cellules phagocytaires du sang
circulant vers le foyer inflammatoire où se trouve la bactérie. Le C5a a également une
puissante activité chimioattractante, c'est-à-dire qu’il attire la bactérie vers le site
inflammatoire.
Il y également d’autres facteurs qui sont dérivés des bactéries comme les formyl-
peptides qui sont de puissants chimioattractants et de puissants activateurs du polynucléaire
neutrophile.
Et enfin les endotoxines, le LPS, les formyl-peptides, le C5a mais également l’agent
pathogène lui-même vont venir activer les macrophages résidants au niveau du foyer
tissulaire. Cette activation va induire la libération d’un certain nombre de facteurs en
particulier : - des facteurs lipidiques : le LTB4 et le PAF
- et un certain nombre de cytokines en particulier des chimiokines comme
l’interleukine 8 qui a un puissant effet sur la migration des polynucléaires
vers le foyer inflammatoire.
Une fois que le polynucléaire se retrouve au niveau du site inflammatoire, il va lui-
même produire un certain nombre de cytokines, l’interleukine 8, qui va permettre le
recrutement d’autres polynucléaires ce qui va amplifier la réponse inflammatoire. Une fois
que les cellules phagocytaires (polynucléaires, monocytes- macrophages) sont présents sur le
site, ils vont phagocyter la bactérie, la dégrader et ensuite les peptides antigéniques issus de
cette dégradation vont être pris en charge par les cellules dendritiques qui vont migrer vers les
organes lymphoïdes, puis elles vont présenter l’Ag aux lymphocytes T et vont ainsi faire le
lien avec la réponse adaptative.
2. le complément :
La cascade du complément va être activé rapidement, comme on l’a vu, par la voie
alterne en une quinzaine de minutes.
Il existe 3 voies majeures d’activation du complément :
la voie classique qui est activée par des complexes Ag-Ac. L’Ag étant, ici, la
membrane d’une bactérie (on est plus dans ce cas là dans l’immunité innée
puisqu’il y a déjà eut production d’anticorps). Le complexe immun active la
sous unité C1q du premier composant du complément, qui va elle-même
activer C1r et C1s.
parallèlement la voie des lectines avec certains carnohydrates des surfaces
microbiennes qui vont se lier à la lectine MBL, qui va activer l’enzyme MASP.
Ces 2 voies vont se retrouver pour activer le C4 puis le C2 et former la C3
convertase classique.
 6
7
8
9
10
11
12
13
6
7
8
9
10
11
12
13
1
/
13
100%