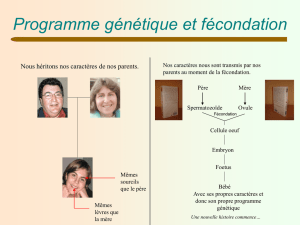
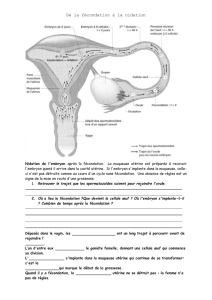
embryologie les oursins

Embryologie 2010 Page 1
EMBRYOLOGIE
LES OURSINS
Les oursins appartiennent au phylum des Echinodermes, il en existe 7000 espèces, toutes
marines, réparties en 5 classes.
- Classe des Echinides (oursins, 900 espèces)
- Classe des Astérides (étoiles des mers)
- Classe des Holothurides (concombres de mer)
- Classe des Ophiurides (ophiures)
- Classe des Crinoïdes (lilas des mers)
I- Anatomie et avantages/inconvénients
1- Anatomie
- Deutérostomiens : formation de l’anus { partir du blastopore (région anale se développe
avant la région buccale) + capacité de régulation des blastomères.
- Segmentation radiaire d’ordre 5 (pentaradiaire) : les plans de clivage sont parallèles ou
perpendiculaires { l’axe primordial (= axe animal-végétatif). Le test calcaire de l’oursin donne
la forme globulaire de l’oursin adulte. Celui-ci est formé de cinq plaques riches en calcium
soudées les unes aux autres 5 doublets de bandes sombres (=région ambulacraire) et claires.
- Segmentation est holoblastique (divisions totales).
- Pas de critères sexuels secondaires qui permettent de
différencier le mâle de la femelle.
- Vie sur fonds benthiques la face orale tournée vers le sol
afin de se nourrir d’algues grâce à un appareil masticateur =
« Lanterne d’Aristote » qui comporte 5 dents calcaires
alternées avec les compas (pièces squelettiques articulées
faisant office de pince) + muscles rétracteurs et transverses
pour faire bouger les dents.
- A l’opposé, on trouve la face aborale, qui constitue la région
ano-génitale qui rejette les excréments.
- Les régions ambulacraires portent les podias.
- Les régions interambulacraires portent les piquants.
- Les piquants ne permettent pas le mouvement, contrairement aux pieds ambulacraires, sur
lesquels sont fixés des podia (= ventouses).
2- Avantages/inconvénients de l’oursin pour l’étude du développement
Avantages :
- Grande production de gamètes : la femelle 107 ovocytes et le mâle 1012 spermatozoïdes,
de façon à augmenter les probabilités de fécondation dans le milieu extérieur (= eau).
- Fécondation et développement externes : très facilement observables et réalisables in vitro.

Embryologie 2010 Page 2
- Le développement de l’embryon d’oursin est très rapide, on obtient au bout de deux jours une
larve et est aussi synchrone, signifiant que toutes les larves croitront de manière identique au
fil du temps.
- L’œuf et l’embryon d’oursin sont transparents (possède peu de réserves = vitellus), ainsi, on
peut facilement observer les types cellulaires { l’intérieur du blastocœle. Il est synchrone :
segmentation à espaces réguliers, chaque embryon se divise au même moment que le voisin. Et
il possède une taille permettant une manipulation facile par micro-injections (ADN, ARN,
morpholinos) grâce à des micropipettes.
- Les embryons sont manipulables facilement, on peut ainsi effectuer aisément des
transplantations.
Protéines et la structure de l’embryon au stade larvaire est simple, avec un faible nombre de
cellules.
Inconvénients :
- Analyse génétique classique impraticable car temps de génération trop long (quelques
années).
- Production saisonnière des gamètes (de janvier à juin pour Paracentrotus lividus).
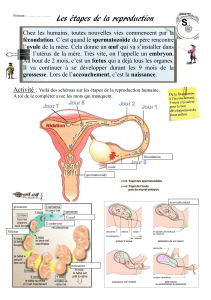
II- La reproduction
Le cycle vital est très long (de la fécondation à la larve pluteus :
2 jours).
Spermatozoïde + ovule
Développement d’une larve nageuse
Echinopluteus (plancton)
Formes rudimentaires et organes larvaires résorbés
Juvénile
Métamorphose et ancrage au sol adulte
1- Les gamètes
Pour les observer, on dissèque les oursins au niveau de
l’équateur.
Les gamètes seront expulsés grâce à des petits canaux
débouchant sur cinq orifices génitaux au niveau de la plaque
aborale. 5 gonades par individus.
Les oursins pratiquant l’oogamie : seul le spermatozoïde est
mobile.
Spz.= un acrosome qui renferme des protéines
(principalement des hydrolases, de l’actine G et de la
bindine). + un flagelle est composé de 9 axonèmes, eux-
mêmes formés de doublets de microtubules ainsi que des
protéines (dynéine interne et externe), permettant aux
microtubules de glisser les uns sur les autres. Il ne possède
qu’une seule membrane.

Embryologie 2010 Page 3
Les ovocytes
Diamètre de 100µm.
Pendant l’ovogenèse, les ovocytes vont maturer et
donner des œufs non fécondés et orientés selon un
axe animal-végétatif : pôle animal et pôle végétatif
respectivement définis par la présence d’un canal
par lequel seront expulsés les deux globules
polaires au cours de la maturation de l’ovocyte, et
par le point d’ancrage de l’œuf { l’ovaire.
L’œuf est lié { l’ovaire pendant l’ovogénèse par le
pôle végétatif.
Il possède 3 types de membranes :
- membrane plasmique (mb interne)
-enveloppe vitelline, collée à la membrane plasmique, forme des microvillosités. Apres la
fécondation, cette mb vitelline va se décoller, pour mettre en place l’enveloppe de fécondation.
- gangue gélatineuse (mb externe) : composée de nombreux sucres, elle reste fine tant que
l’œuf se trouve dans l’ovaire, et servira par la suite de flotteur { l’œuf (se gorge d’eau une fois
l’œuf pondu). Elle va devenir très épaisse et va jouer un rôle de reconnaissance du
spermatozoïde durant la fécondation : elle est capable d’attirer via un mécanisme de
chimiotactisme les spermatozoïdes d’oursins de même espèce. Impliqué dans ce mécanisme,
on a pu isoler un peptide de 14 acides aminés, la resact, synthétisée par la gangue gélatineuse.
Sous la membrane plasmique, on trouve une grande quantité de vésicules, les granules
corticaux. Ceux-ci auront un rôle important à jouer lors de la fécondation.
Après expulsion, le courant va disséminer les gamètes dans le milieu marin, et ils vont se
rencontrer pour faire la fécondation. Une espèce d’oursins sera incapable de féconder une
espèce étrangère, du fait de mécanismes propres à leurs gamètes respectifs.
2- La fécondation
Les oursins n’ont pas de caractères sexuels secondaires mais si un oursin possède de
nombreux gamètes matures prêts à être expulsés, il y aura en son sein, soit une substance
orangée, soit une substance blanche près du pôle aboral. Les ovocytes et les œufs sont très
orangés, car ils expriment un pigment au niveau de leur cortex.
Les œufs sont beaucoup plus gros et denses que les spermatozoïdes, les oursins étant
anisogames.
Contrairement { d’autres espèces animales, le spermatozoïde d’oursin va pouvoir pénétrer
dans l’œuf { n’importe quel niveau. Le premier contact entre l’œuf et les spermatozoïdes va
activer les deux gamètes :
Réaction acrosomale du spermatozoïde
= exocytose de la vésicule acrosomale : les protéines vont être libérées et les hydrolases qui
vont digérer la gangue gélatineuse. L’actine sous forme globulaire (G) va polymériser en actine
F et former le filament acrosomal.
La bindine de la membrane de l’acrosome va être reconnue par un récepteur au niveau de la
membrane vitelline de l’œuf.

Embryologie 2010 Page 4
La création du complexe bindine/récepteur permet la fusion des deux membranes, et la
pénétration du noyau dans l’œuf et permet d’éviter une fécondation interespèce. Une enzyme
va ensuite digérer la mb vitelline, et la tête du spermatozoïde va fusionner avec la membrane
plasmique de l’œuf.
2 réactions de l’œuf pour éviter la polyspermie
L’œuf est chargé négativement avant la fécondation (-75
mV), en cas de fécondation, un flux entrant ou sortant d’ions va
bouleverser ce potentiel. La dépolarisation induite empêchera
la fixation des spermatozoïdes surnuméraires sur l’œuf. La
réponse à la fécondation est rapide, et le potentiel de
membrane retrouve un niveau normal au bout d’une dizaine de
minutes.
Réaction corticale : les granules corticaux vont déverser leur contenu protéique par
exocytose, ce qui va provoquer un soulèvement de la membrane vitelline : elle va se décoller de
la membrane plasmique. En effet, des protéases vont couper les liens entre les deux
membranes. Des sucres vont être relargués et créer un gradient osmotique pour permettre à
l’eau de mer de rentrer entre les deux membranes, et créer l’espace périvitellin.
L’enveloppe de fécondation est la membrane vitelline qui a été modifiée après la fécondation.
C’est une enveloppe très résistante, qui assure une protection définitive contre la polyspermie.
Une couche de hyaline va entourer l’œuf pour assurer la cohésion des cellules pendant la
division.
Schéma de l’œuf fécondé

Embryologie 2010 Page 5
III- De L’œuf à l’oursin
1- La segmentation
Après une demi-heure, il y a fusion des noyaux des deux gamètes, et formation d’un zygote. On
appelle cette fusion amphimixie. L’œuf va ensuite se diviser et mener à un organisme
pluricellulaire par une segmentation orientée selon l’axe animal-végétatif, holoblastique (d’un
pôle { l’autre), totale, radiaire et réductive (les cellules ont des tailles de plus en plus petites
mais la taille de l’embryon
reste la même).
Le 1er plan de clivage
est méridien, débute sous
forme d’une contraction
corticale. Les 2 cellules issues
de cette segmentation sont
des blastomères.
Stade 2 : Le 2e plan
clivage, perpendiculaire au
premier, va donner naissance
à 4 blastomères de taille
identique.
Stade 4 : Le 3e plan de clivage va être situé { l’équateur, et donner 4 cellules dans
l’hémisphère du pôle animal, et 4 cellules dans l’hémisphère du pôle végétal. La bande
pigmentaire est contenue dans les blastomères inférieurs
Stade 8 : Les 4 cellules du pôle végétal vont se diviser asymétriquement avec formation
de petites cellules au pôle végétatif, et de grandes cellules au pôle animal + formation d’une
cavité entre les blastomères = blastocœle.
Stade 16 : les blastomères supérieurs vont se diviser de façon méridienne et donneront
les mésomères. Les blastomères inférieurs vont se diviser de façon latitudinale décalée et
donneront 4 macromères et 4 micromères.
Stade 32 : Les mésomères subissent une division équatoriale et donneront les cellules
an1 et an2. Les micromères vont subir une division latitudinale asymétrique pour donner
naissance aux grands et aux petits micromères. Les mésomères se diviseront de façon
méridienne pour en former 8.
Stade 64 : les macromères subissent une division équatoriale pour donner deux assises
végétatives veg1 et veg2
A partir du stade 128 cellules on obtient un embryon appelé blastula.
Chaque cellule va acquérir un cil, servant à faire tourner la blastula. A la fin de ce stade, il va y
avoir éclosion de l’enveloppe de fécondation (10-12h après la fécondation): les cellules de la
partie animale vont synthétiser l’enzyme d’éclosion qui va digérer l’enveloppe et libérer la
blastula flotteuse capable de « nager » grâce à ses cils.
Stade 4
Stade 64
 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%