Microbiologie Générale : Classification et Structure Bactérienne
Telechargé par
Issam Laaouach

See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/339461316
microbiologie générale Dr TOUAITIA
Chapter · February 2020
CITATIONS
0
READS
817
1 author:
Some of the authors of this publication are also working on these related projects:
antibiorésistance View project
antibiorésistance bactérienne View project
Touaitia Rahima
Badji Mokhtar - Annaba University
7 PUBLICATIONS4 CITATIONS
SEE PROFILE
All content following this page was uploaded by Touaitia Rahima on 24 February 2020.
The user has requested enhancement of the downloaded file.

Microbiologie générale Dr. Touaitia R
1
INTRODUCTION :
La microbiologie a souvent été définie comme l’étude d’organismes trop petits pour
être vus à l’œil nu c’est-à-dire l’étude des microorganismes.
Du fait que les objets d’une taille inférieure à environ un millimètre de diamètre ne
peuvent être clairement vus à l’œil nu et doivent être examinés à l’aide d’un
microscope, la microbiologie est principalement concernée par des organismes et des
agents aussi petits ou plus petits. Cependant, certains microorganismes, en particulier
certains micro-organismes eucaryotes, sont visibles sans microscope, ainsi les
moisissures du pain et les algues filamenteuses.
1. Place des micro-organismes dans le monde vivant :
Depuis leur découverte par Anthony van Leeuwenhoeck, la place des bactéries dans le
monde vivant a beaucoup évoluée. Le botaniste suédois Carl van Linné (1735),
élabora une première classification des organismes vivants en deux règnes Plantae et
Animalia. En 1857, Karl van Nageli proposa de classer les bactéries et les
champignons dans le règne des Plantes.
2. Classification de Haeckel
En 1866, E. Haeckel divise le monde vivant en trois règnes, le règne animal, le règne
végétal et le règne des protistes qui rassemble les algues, les protozoaires, les
champignons et les bactéries.
3. Les cellules eucaryotes et procaryotes selon Edward Chatton
En 1937 et grâce à l’invention du microscope électronique, Edward Chatton mis en
opposition deux types de cellules, la cellule eucaryote et la cellule procaryote.
En 1938, H.F. Copeland sépare le règne des bactéries (ou "Monera") de celui des
protistes. Cette définition des procaryotes fut renforcée en 1961 par Roger Stanier.

Microbiologie générale Dr. Touaitia R
2
Tableau I: Classification des êtres vivants
Principaux
Groupes
Bactéries
Algues
protozoaires
Levures
Moisissures
Champignons
Animaux
Végétaux
Type
Cellulaire
Procaryotes
Eucaryotes
Eucaryotes
Eucaryotes
Eucaryotes
Organisation
Biologique
Unicellulaires
Unicellulaires
ou
Pluricellulaires
(algues)
Unicellulaires
ou
Pluricellulaires
Pluricellulaires
Pluricellulaires
Taille
Microscopique
Microscopique
ou
Macroscopique
Microscopique
ou
Macroscopique
Macroscopique
Macroscopique
4. Distinction entre cellule eucaryote et cellule procaryote :
Le tableau suivant résume les caractéristiques des eucaryotes et des procaryotes tout
en faisant apparaitre les caractères de différenciation.
Caractéristiques
Cellule Procaryote
Cellule Eucaryote
Taille typique
1-10 µm
10-100 µm
Type de noyeau
nucléoïde (pas de véritable
noyau)
vrai noyau avec double
membrane
Division de la cellule
division
simple
mitose (réplication de la cellule)
méiose (menant à la formation
de gamètes)
Organisation génétique
Membrane nucléaire
Non
oui
Nombre de chromosomes
Chromosome circulaire
Histones
1 chromosome (Haploïde)
Oui
Non
Plusieurs (Diploïde)
Non
oui
Structures cellulaires et organites
Réticulum endoplasmique
Non
oui
Appareil de Golgi
Non
oui
Mitochondries
Non
oui
Paroi cellulaire avec
peptidoglycane
Oui
non

Microbiologie générale Dr. Touaitia R
3
LES BACTERIES :
Les bactéries sont des micro-organismes que l’on rencontre pratiquement partout.
Leur présence est souvent manifeste : les blessures s’infectent, le lait surit, la viande
se putréfie, mais, on ne peut les voir qu’au microscope.
1. Formes et groupements :
Elles peuvent être observées :
soit à l’état frais, entre lame et lamelle, à partir d’un étalement de liquide
biologique (pus, crachats,…) ou d’une culture en milieu liquide.
soit, après coloration, à partir d’un étalement sur une lame, séché et fixé.
La forme des bactéries permet de les classer en :
Bactéries sphériques: Cocci
Les cocci peuvent se grouper en :
Diplocoques tétrades en chaînettes en grappes
Bactéries cylindriques : bacilles
Les bacilles peuvent se grouper en :
Bacilles isolés Diplobacilles
Microbiologie générale Dr. Touaitia R
3
LES BACTERIES :
Les bactéries sont des micro-organismes que l’on rencontre pratiquement partout.
Leur présence est souvent manifeste : les blessures s’infectent, le lait surit, la viande
se putréfie, mais, on ne peut les voir qu’au microscope.
1. Formes et groupements :
Elles peuvent être observées :
soit à l’état frais, entre lame et lamelle, à partir d’un étalement de liquide
biologique (pus, crachats,…) ou d’une culture en milieu liquide.
soit, après coloration, à partir d’un étalement sur une lame, séché et fixé.
La forme des bactéries permet de les classer en :
Bactéries sphériques: Cocci
Les cocci peuvent se grouper en :
Diplocoques tétrades en chaînettes en grappes
Bactéries cylindriques : bacilles
Les bacilles peuvent se grouper en :
Bacilles isolés Diplobacilles
Microbiologie générale Dr. Touaitia R
3
LES BACTERIES :
Les bactéries sont des micro-organismes que l’on rencontre pratiquement partout.
Leur présence est souvent manifeste : les blessures s’infectent, le lait surit, la viande
se putréfie, mais, on ne peut les voir qu’au microscope.
1. Formes et groupements :
Elles peuvent être observées :
soit à l’état frais, entre lame et lamelle, à partir d’un étalement de liquide
biologique (pus, crachats,…) ou d’une culture en milieu liquide.
soit, après coloration, à partir d’un étalement sur une lame, séché et fixé.
La forme des bactéries permet de les classer en :
Bactéries sphériques: Cocci
Les cocci peuvent se grouper en :
Diplocoques tétrades en chaînettes en grappes
Bactéries cylindriques : bacilles
Les bacilles peuvent se grouper en :
Bacilles isolés Diplobacilles

Microbiologie générale Dr. Touaitia R
4
Streptobacilles Bacilles incurvés
Bactéries spiralées :
Hélicoïdales : Tréponèmes Ramifiées : Actinomycètes
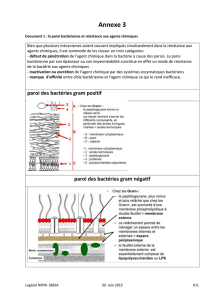
2. Structure de la cellule bactérienne :
Voici la cellule bactérienne et ses principales composantes : la capsule, paroi
cellulaire, membrane plasmique, cytoplasme, flagelle, et nucléoide.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
1
/
36
100%