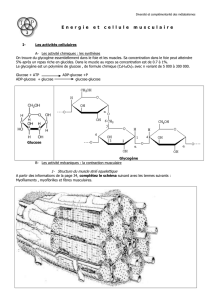
•INTRODUCTION
•-Illustration de l interdépendance des voies métaboliques
•-L'anabolisme
•-Le catabolisme
•LA GLYCOLYSE
•-La phase préparatoire
•-La phase de remboursement
•-Le rendement énergétique de la conversion du glucose en pyruvate
•-Entrée des oses dans la glycolyse
•-Régulation de la glycolyse
•LA NEOGLUCOGENESE
•-La biotine est un transporteur mobile de CO2 activé
•-La pyruvate carboxylase est activée par l’acétyl CoA
•-L’oxaloacétate revient au cytosol par une navette et est transformé en
phosphoénolpyruvate
•-Six liaisons phosphate riches en énergie sont dépensées au cours de la synthèse
du glucose à partir du pyruvate
•-La néoglucogenèse et la glycolyse sont régulées réciproquement

•LA NAVETTE MALATE-ASPARTATE
•LA NAVETTE DU GLYCEROL-PHOSPHATE
•LE CYCLE DE L’ACIDE CITRIQUE
•-Formation de l’acétyl CoA à partir du pyruvate
•-L’oxaloacétate se condense avec l’acétyl coenzymeA pour former du citrate
•-Le citrate est isomérisé en isocitrate
•-L’isocitrate est oxydé et décarboxylé en a-cétoglutarate
•-Le succinyl coenzyme A est formé par la décarboxylation oxydative de l’a-
cétoglutarate
•-Une liaison phosphate riche en énergie est formée à partir du succinyl coenzyme
A
•-La régénération de l’oxaloacétate par l’oxydation du succinate
•-Le contrôle du cycle de krebs
•-Le cycle du glyoxylate
•LA CHAINE RESPIRATOIRE ET PHOSPHORYLATION OXYDATIVE
•-La chaine d’oxydoréduction
•-Oxydoréductions
•-Les flux de protons, libération d’énergie osmotique
•-La phosphorylation
•-Le transport de l’ATP
•-Régulation de la chaine respiratoire

•FERMENTATIONS ALCOLIQUE ET LACTIQUE
•LE PYRUVATE PEUT ETRE TRANSFORME EN ETHANOL, LACTATE OU ACETYL
COENZYME A
•LE METABOLISME DU GLYCOGENE
•-La phosphorylase catalyse le clivage phosphorylitique du glycogène en glucose
1-phosphate.
•-Un enzyme débranchant est également nécessaire à la destruction du glycogène.
•-La phosphoglucomutase transforme le glucose 1-phosphate en glucose 6-
phosphate.
•-Le foie contient de la glucose 6-phosphatase, un enzyme hydrolytique absent du
muscle.
•-Le glycogène et synthétisé et dégradé par différentes voies.
•-La glycogène synthase catalyse le transfert du glucose de l’UDP-glucose à la
chaîne en croissance.
•-Un enzyme branchant forme les liaisons a-1,6.
•-Contrôle de la synthèse et de la dégradation du glycogène
•

•LA VOIE DES PENTOSES PHOSPHATE
•-Deux NADPH sont formés au cours de la conversion du glucose 6-phosphate en
ribulose 5-phosphate
•-Le ribulose 5-phoshate est isomérisé en ribose 5-phosphate par l‘intermédiaire
d’un ènediol.
•-La voie des pentoses phosphate et la glycolyse sont reliées par la transcétolase
et la transaldolase.
•-La vitesse de la voie des pentoses phosphate est contrôlée par le niveau de
NADP+.
•METABOLISME DES ACIDES GRAS
•-Les acides gras sont dégradés par l’élimination séquentielle de deux unités
carbonées
•-Mécanisme de l’oxydation des A.G saturés.
•-Oxydation des AG insaturés :
•-Biosynthèse des A.G
•-Régulation de la synthèse des acides gras
•-Elongation des AG à longue chaîne et désaturation

•BIOSYNTHESE DU CHOLESTEROL
•-Le bilan de la synthèse des stérols
•-Régulation de la synthèse du cholesterol
•METABOLISME DES ACIDES AMINES
•-Proteolyse
•-Transamination
•-Désamination
•- Transamination et désamination oxydative du glutamate
•-Cycle de l’urée
•-Lien entre le cycle de l’urée et le cycle de l‘acide citrique
•-Devenir du squelette de carbone
•-Biosynthèse des acides amines
•- Fixation de l’azote moléculaire
•LES CORPS CETONIQUES
•METABOLISME DES ACIDES NUCLEIQUES
•-Biosynthèse des nucléotides
•-Catabolisme des acides nucléiques
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
1
/
222
100%