L'épidémie de Extended-Spectrum- Lactamase-ProducingL'épidémie de Extended-Spectrum- Lactamase-Producing
Escherichia coli
ST131 est conduite par un seul sous-clone hautement pathogène,
H
30-Rx
Escherichia coli
ST131 est conduite par un seul sous-clone hautement pathogène,
H
30-Rx
Escherichia coli
ST131 est conduite par un seul sous-clone hautement pathogène,
H
30-Rx
Escherichia coli
ST131 est conduite par un seul sous-clone hautement pathogène,
H
30-Rx
Lance B. Prix, un B James R. Johnson, c Maliha Aziz, un B Connie Clabots, c Brian Johnston, c Veronika Tchesnokova, ré Lora Nordstrom, bLance B. Prix, un B James R. Johnson, c Maliha Aziz, un B Connie Clabots, c Brian Johnston, c Veronika Tchesnokova, ré Lora Nordstrom, bLance B. Prix, un B James R. Johnson, c Maliha Aziz, un B Connie Clabots, c Brian Johnston, c Veronika Tchesnokova, ré Lora Nordstrom, bLance B. Prix, un B James R. Johnson, c Maliha Aziz, un B Connie Clabots, c Brian Johnston, c Veronika Tchesnokova, ré Lora Nordstrom, bLance B. Prix, un B James R. Johnson, c Maliha Aziz, un B Connie Clabots, c Brian Johnston, c Veronika Tchesnokova, ré Lora Nordstrom, bLance B. Prix, un B James R. Johnson, c Maliha Aziz, un B Connie Clabots, c Brian Johnston, c Veronika Tchesnokova, ré Lora Nordstrom, bLance B. Prix, un B James R. Johnson, c Maliha Aziz, un B Connie Clabots, c Brian Johnston, c Veronika Tchesnokova, ré Lora Nordstrom, bLance B. Prix, un B James R. Johnson, c Maliha Aziz, un B Connie Clabots, c Brian Johnston, c Veronika Tchesnokova, ré Lora Nordstrom, bLance B. Prix, un B James R. Johnson, c Maliha Aziz, un B Connie Clabots, c Brian Johnston, c Veronika Tchesnokova, ré Lora Nordstrom, bLance B. Prix, un B James R. Johnson, c Maliha Aziz, un B Connie Clabots, c Brian Johnston, c Veronika Tchesnokova, ré Lora Nordstrom, bLance B. Prix, un B James R. Johnson, c Maliha Aziz, un B Connie Clabots, c Brian Johnston, c Veronika Tchesnokova, ré Lora Nordstrom, bLance B. Prix, un B James R. Johnson, c Maliha Aziz, un B Connie Clabots, c Brian Johnston, c Veronika Tchesnokova, ré Lora Nordstrom, bLance B. Prix, un B James R. Johnson, c Maliha Aziz, un B Connie Clabots, c Brian Johnston, c Veronika Tchesnokova, ré Lora Nordstrom, bLance B. Prix, un B James R. Johnson, c Maliha Aziz, un B Connie Clabots, c Brian Johnston, c Veronika Tchesnokova, ré Lora Nordstrom, b
Maria Billig, ré Sujay Chattopadhyay, ré Marc Stegger, a, e Paal S. Andersen, a, e Talima Pearson, F Kim Riddell, g Peggy Rogers, gMaria Billig, ré Sujay Chattopadhyay, ré Marc Stegger, a, e Paal S. Andersen, a, e Talima Pearson, F Kim Riddell, g Peggy Rogers, gMaria Billig, ré Sujay Chattopadhyay, ré Marc Stegger, a, e Paal S. Andersen, a, e Talima Pearson, F Kim Riddell, g Peggy Rogers, gMaria Billig, ré Sujay Chattopadhyay, ré Marc Stegger, a, e Paal S. Andersen, a, e Talima Pearson, F Kim Riddell, g Peggy Rogers, gMaria Billig, ré Sujay Chattopadhyay, ré Marc Stegger, a, e Paal S. Andersen, a, e Talima Pearson, F Kim Riddell, g Peggy Rogers, gMaria Billig, ré Sujay Chattopadhyay, ré Marc Stegger, a, e Paal S. Andersen, a, e Talima Pearson, F Kim Riddell, g Peggy Rogers, gMaria Billig, ré Sujay Chattopadhyay, ré Marc Stegger, a, e Paal S. Andersen, a, e Talima Pearson, F Kim Riddell, g Peggy Rogers, gMaria Billig, ré Sujay Chattopadhyay, ré Marc Stegger, a, e Paal S. Andersen, a, e Talima Pearson, F Kim Riddell, g Peggy Rogers, gMaria Billig, ré Sujay Chattopadhyay, ré Marc Stegger, a, e Paal S. Andersen, a, e Talima Pearson, F Kim Riddell, g Peggy Rogers, gMaria Billig, ré Sujay Chattopadhyay, ré Marc Stegger, a, e Paal S. Andersen, a, e Talima Pearson, F Kim Riddell, g Peggy Rogers, gMaria Billig, ré Sujay Chattopadhyay, ré Marc Stegger, a, e Paal S. Andersen, a, e Talima Pearson, F Kim Riddell, g Peggy Rogers, gMaria Billig, ré Sujay Chattopadhyay, ré Marc Stegger, a, e Paal S. Andersen, a, e Talima Pearson, F Kim Riddell, g Peggy Rogers, gMaria Billig, ré Sujay Chattopadhyay, ré Marc Stegger, a, e Paal S. Andersen, a, e Talima Pearson, F Kim Riddell, g Peggy Rogers, gMaria Billig, ré Sujay Chattopadhyay, ré Marc Stegger, a, e Paal S. Andersen, a, e Talima Pearson, F Kim Riddell, g Peggy Rogers, g
Delia Scholes, h Barbara Kahl, je Paul Keim, un F Evgeni V. Sokurenko réDelia Scholes, h Barbara Kahl, je Paul Keim, un F Evgeni V. Sokurenko réDelia Scholes, h Barbara Kahl, je Paul Keim, un F Evgeni V. Sokurenko réDelia Scholes, h Barbara Kahl, je Paul Keim, un F Evgeni V. Sokurenko réDelia Scholes, h Barbara Kahl, je Paul Keim, un F Evgeni V. Sokurenko réDelia Scholes, h Barbara Kahl, je Paul Keim, un F Evgeni V. Sokurenko réDelia Scholes, h Barbara Kahl, je Paul Keim, un F Evgeni V. Sokurenko réDelia Scholes, h Barbara Kahl, je Paul Keim, un F Evgeni V. Sokurenko ré
Division de la génomique Pathogen, l'Institut de recherche génomique translationnelle, Flagstaff, Arizona, États-Unis une; Ministère de la santé et l'environnement, l'Université George Washington, Washington, DC, États-Unis b; Veterans Division de la génomique Pathogen, l'Institut de recherche génomique translationnelle, Flagstaff, Arizona, États-Unis une; Ministère de la santé et l'environnement, l'Université George Washington, Washington, DC, États-Unis b; Veterans Division de la génomique Pathogen, l'Institut de recherche génomique translationnelle, Flagstaff, Arizona, États-Unis une; Ministère de la santé et l'environnement, l'Université George Washington, Washington, DC, États-Unis b; Veterans Division de la génomique Pathogen, l'Institut de recherche génomique translationnelle, Flagstaff, Arizona, États-Unis une; Ministère de la santé et l'environnement, l'Université George Washington, Washington, DC, États-Unis b; Veterans Division de la génomique Pathogen, l'Institut de recherche génomique translationnelle, Flagstaff, Arizona, États-Unis une; Ministère de la santé et l'environnement, l'Université George Washington, Washington, DC, États-Unis b; Veterans
Affairs Medical Center et de l'Université du Minnesota, Minneapolis, Minnesota, États-Unis c; Département de microbiologie, Université de Washington School of Medicine, Seattle, Washington, États-Unis ré; Contrôle de la Affairs Medical Center et de l'Université du Minnesota, Minneapolis, Minnesota, États-Unis c; Département de microbiologie, Université de Washington School of Medicine, Seattle, Washington, États-Unis ré; Contrôle de la Affairs Medical Center et de l'Université du Minnesota, Minneapolis, Minnesota, États-Unis c; Département de microbiologie, Université de Washington School of Medicine, Seattle, Washington, États-Unis ré; Contrôle de la Affairs Medical Center et de l'Université du Minnesota, Minneapolis, Minnesota, États-Unis c; Département de microbiologie, Université de Washington School of Medicine, Seattle, Washington, États-Unis ré; Contrôle de la Affairs Medical Center et de l'Université du Minnesota, Minneapolis, Minnesota, États-Unis c; Département de microbiologie, Université de Washington School of Medicine, Seattle, Washington, États-Unis ré; Contrôle de la
microbiologie et l'infection, Statens Serum Institut, Copenhague, Danemark e; Centre de Génétique et génomique microbienne, Northern Arizona University, Flagstaff, Arizona, États-Unis F; Groupe Santé Laboratoire clinique, Group microbiologie et l'infection, Statens Serum Institut, Copenhague, Danemark e; Centre de Génétique et génomique microbienne, Northern Arizona University, Flagstaff, Arizona, États-Unis F; Groupe Santé Laboratoire clinique, Group microbiologie et l'infection, Statens Serum Institut, Copenhague, Danemark e; Centre de Génétique et génomique microbienne, Northern Arizona University, Flagstaff, Arizona, États-Unis F; Groupe Santé Laboratoire clinique, Group microbiologie et l'infection, Statens Serum Institut, Copenhague, Danemark e; Centre de Génétique et génomique microbienne, Northern Arizona University, Flagstaff, Arizona, États-Unis F; Groupe Santé Laboratoire clinique, Group microbiologie et l'infection, Statens Serum Institut, Copenhague, Danemark e; Centre de Génétique et génomique microbienne, Northern Arizona University, Flagstaff, Arizona, États-Unis F; Groupe Santé Laboratoire clinique, Group
Health Cooperative, Seattle, Washington, États-Unis g; Institut de recherche sur la santé du groupe, Group Health Cooperative, Seattle, Washington, États-Unis h; Institut de microbiologie médicale, UniversitätsklinikumMunster, Health Cooperative, Seattle, Washington, États-Unis g; Institut de recherche sur la santé du groupe, Group Health Cooperative, Seattle, Washington, États-Unis h; Institut de microbiologie médicale, UniversitätsklinikumMunster, Health Cooperative, Seattle, Washington, États-Unis g; Institut de recherche sur la santé du groupe, Group Health Cooperative, Seattle, Washington, États-Unis h; Institut de microbiologie médicale, UniversitätsklinikumMunster, Health Cooperative, Seattle, Washington, États-Unis g; Institut de recherche sur la santé du groupe, Group Health Cooperative, Seattle, Washington, États-Unis h; Institut de microbiologie médicale, UniversitätsklinikumMunster, Health Cooperative, Seattle, Washington, États-Unis g; Institut de recherche sur la santé du groupe, Group Health Cooperative, Seattle, Washington, États-Unis h; Institut de microbiologie médicale, UniversitätsklinikumMunster,
Münster, Allemagne jeMünster, Allemagne je
LBP et JRJ ont contribué également au projet.
ABSTRAIT le
Escherichia coli
Type de séquence 131 clone (ST131) est bien connu pour les infections extra-intestinales, de la résistance fl uoroquinolone et la production ABSTRAIT le
Escherichia coli
Type de séquence 131 clone (ST131) est bien connu pour les infections extra-intestinales, de la résistance fl uoroquinolone et la production ABSTRAIT le
Escherichia coli
Type de séquence 131 clone (ST131) est bien connu pour les infections extra-intestinales, de la résistance fl uoroquinolone et la production ABSTRAIT le
Escherichia coli
Type de séquence 131 clone (ST131) est bien connu pour les infections extra-intestinales, de la résistance fl uoroquinolone et la production
spectrumbeta-lactamase étendu (ESBL), imputables à un CTX-M-15 codant pour un élément mobile. Ici, nous avons appliqué fi eld gel puisée électrophorèse (PFGE) et le
séquençage du génome entier pour reconstituer l'histoire évolutive du clone ST131. analyses cluster basé PFGE a suggéré que les deux fl résistance à la uoroquinolone et la
production BLSE avaient été acquises par plusieurs sous-lignées de ST131 à travers des événements génétiques indépendants. En revanche, l'ensemble-genomesequence basée
sur l'analyse phylogénomique plus robuste a révélé que la résistance fl uoroquinolone a été con fi nie presque entièrement à un seul, en expansion rapide ST131 sous-clone,
désigné
H
30-R. Frappant de constater que, 91% des isolats produisant CTX-M-15 a également appartenu à un seul, bien dé fi ni clade imbriqué dans
H
30-R, qui a été nommé
H
30-Rx désigné
H
30-R. Frappant de constater que, 91% des isolats produisant CTX-M-15 a également appartenu à un seul, bien dé fi ni clade imbriqué dans
H
30-R, qui a été nommé
H
30-Rx désigné
H
30-R. Frappant de constater que, 91% des isolats produisant CTX-M-15 a également appartenu à un seul, bien dé fi ni clade imbriqué dans
H
30-R, qui a été nommé
H
30-Rx désigné
H
30-R. Frappant de constater que, 91% des isolats produisant CTX-M-15 a également appartenu à un seul, bien dé fi ni clade imbriqué dans
H
30-R, qui a été nommé
H
30-Rx désigné
H
30-R. Frappant de constater que, 91% des isolats produisant CTX-M-15 a également appartenu à un seul, bien dé fi ni clade imbriqué dans
H
30-R, qui a été nommé
H
30-Rx désigné
H
30-R. Frappant de constater que, 91% des isolats produisant CTX-M-15 a également appartenu à un seul, bien dé fi ni clade imbriqué dans
H
30-R, qui a été nommé
H
30-Rx désigné
H
30-R. Frappant de constater que, 91% des isolats produisant CTX-M-15 a également appartenu à un seul, bien dé fi ni clade imbriqué dans
H
30-R, qui a été nommé
H
30-Rx
en raison de sa résistance plus étendue. En dépit de sa relation étroite avec clonale
H
30Rx, l'organe mobile CTX-M-15 a été inséré dans le plasmide de façon variable et les en raison de sa résistance plus étendue. En dépit de sa relation étroite avec clonale
H
30Rx, l'organe mobile CTX-M-15 a été inséré dans le plasmide de façon variable et les en raison de sa résistance plus étendue. En dépit de sa relation étroite avec clonale
H
30Rx, l'organe mobile CTX-M-15 a été inséré dans le plasmide de façon variable et les
emplacements chromosomiques au sein de la
H
génome 30-Rx. Le dépistage d'une grande collection de clinique récente
E. coli
isole les deux con fi rmé l'expansion mondiale clonale de
H
génome 30-Rx. Le dépistage d'une grande collection de clinique récente
E. coli
isole les deux con fi rmé l'expansion mondiale clonale de
H
génome 30-Rx. Le dépistage d'une grande collection de clinique récente
E. coli
isole les deux con fi rmé l'expansion mondiale clonale de
H
génome 30-Rx. Le dépistage d'une grande collection de clinique récente
E. coli
isole les deux con fi rmé l'expansion mondiale clonale de
H
30-Rx et a révélé son association disproportionnée avec la septicémie (risque relatif, 7,5;
P <
0,001). Ensemble, ces résultats suggèrent que la forte prévalence de la
H
30-Rx et a révélé son association disproportionnée avec la septicémie (risque relatif, 7,5;
P <
0,001). Ensemble, ces résultats suggèrent que la forte prévalence de la
H
30-Rx et a révélé son association disproportionnée avec la septicémie (risque relatif, 7,5;
P <
0,001). Ensemble, ces résultats suggèrent que la forte prévalence de la
H
30-Rx et a révélé son association disproportionnée avec la septicémie (risque relatif, 7,5;
P <
0,001). Ensemble, ces résultats suggèrent que la forte prévalence de la
production CTX-M-15 parmi les isolats ST131 est principalement attribuable à l'expansion d'un sous-clone unique, très virulent,
H
30-Rx.production CTX-M-15 parmi les isolats ST131 est principalement attribuable à l'expansion d'un sous-clone unique, très virulent,
H
30-Rx.production CTX-M-15 parmi les isolats ST131 est principalement attribuable à l'expansion d'un sous-clone unique, très virulent,
H
30-Rx.
IMPORTANCE Nous avons appliqué une approche génomique avancée pour étudier l'histoire de l'évolution récente de l'un des plus importantsIMPORTANCE Nous avons appliqué une approche génomique avancée pour étudier l'histoire de l'évolution récente de l'un des plus importants
Escherichia coli
les souches en circulation aujourd'hui. Cette souche, appelée type de séquence 131 (ST131), provoque la vessie multirésistante, les reins et les infections de la circulation
Escherichia coli
les souches en circulation aujourd'hui. Cette souche, appelée type de séquence 131 (ST131), provoque la vessie multirésistante, les reins et les infections de la circulation
sanguine dans le monde entier. La prévalence croissante de la résistance aux antibiotiques dans
E. coli
rend ces infections plus dif fi ciles à traiter et est conduit à increasedmortality. Des sanguine dans le monde entier. La prévalence croissante de la résistance aux antibiotiques dans
E. coli
rend ces infections plus dif fi ciles à traiter et est conduit à increasedmortality. Des sanguine dans le monde entier. La prévalence croissante de la résistance aux antibiotiques dans
E. coli
rend ces infections plus dif fi ciles à traiter et est conduit à increasedmortality. Des
études antérieures ont suggéré que de nombreuses souches différentes de ST131 ont gagné la résistance aux céphalosporines à spectre étendu de manière indépendante. En revanche, nos
recherches indiquent que la plupart des souches extendedspectrum-résistante aux céphalosporines ST131 appartiennent à un seul sous-clone hautement pathogène, appelé
H
30-Rx. La nature recherches indiquent que la plupart des souches extendedspectrum-résistante aux céphalosporines ST131 appartiennent à un seul sous-clone hautement pathogène, appelé
H
30-Rx. La nature recherches indiquent que la plupart des souches extendedspectrum-résistante aux céphalosporines ST131 appartiennent à un seul sous-clone hautement pathogène, appelé
H
30-Rx. La nature
clonale de
H
30-Rx peut offrir des possibilités de stratégies de contrôle axées sur la prévention vaccins ou de transmission, ce qui pourrait prendre de l'importance que
H
30 Rx et autres clonale de
H
30-Rx peut offrir des possibilités de stratégies de contrôle axées sur la prévention vaccins ou de transmission, ce qui pourrait prendre de l'importance que
H
30 Rx et autres clonale de
H
30-Rx peut offrir des possibilités de stratégies de contrôle axées sur la prévention vaccins ou de transmission, ce qui pourrait prendre de l'importance que
H
30 Rx et autres clonale de
H
30-Rx peut offrir des possibilités de stratégies de contrôle axées sur la prévention vaccins ou de transmission, ce qui pourrait prendre de l'importance que
H
30 Rx et autres clonale de
H
30-Rx peut offrir des possibilités de stratégies de contrôle axées sur la prévention vaccins ou de transmission, ce qui pourrait prendre de l'importance que
H
30 Rx et autres
pathogènes extraintestinal
E. coli
sous-clones deviennent résistantes à nos antibiotiques.pathogènes extraintestinal
E. coli
sous-clones deviennent résistantes à nos antibiotiques.pathogènes extraintestinal
E. coli
sous-clones deviennent résistantes à nos antibiotiques.
Reçu 22 mai 2013 Accepté 12 Novembre 2013 publié 17 Décembre 2013Reçu 22 mai 2013 Accepté 12 Novembre 2013 publié 17 Décembre 2013Reçu 22 mai 2013 Accepté 12 Novembre 2013 publié 17 Décembre 2013Reçu 22 mai 2013 Accepté 12 Novembre 2013 publié 17 Décembre 2013Reçu 22 mai 2013 Accepté 12 Novembre 2013 publié 17 Décembre 2013Reçu 22 mai 2013 Accepté 12 Novembre 2013 publié 17 Décembre 2013
Citation Prix LB, Johnson JR, Aziz M, Clabots C, Johnston B, Tchesnokova V, Nordstrom L, Billig M, Chattopadhyay S, Stegger M, Andersen PS, Pearson T, Riddell K, Rogers P, Scholes D, Kahl B, Keim P , Sokurenko EV. 2013. Citation Prix LB, Johnson JR, Aziz M, Clabots C, Johnston B, Tchesnokova V, Nordstrom L, Billig M, Chattopadhyay S, Stegger M, Andersen PS, Pearson T, Riddell K, Rogers P, Scholes D, Kahl B, Keim P , Sokurenko EV. 2013.
L'épidémie de prolongée spectrum- lactamase productrices
Escherichia coli
ST131 est entraîné par un seul sous-clone hautement pathogène,
H
30-Rx. MBIO 4 (6): e00377-13. doi: 10.1128 / mBio.00377-13.L'épidémie de prolongée spectrum- lactamase productrices
Escherichia coli
ST131 est entraîné par un seul sous-clone hautement pathogène,
H
30-Rx. MBIO 4 (6): e00377-13. doi: 10.1128 / mBio.00377-13.L'épidémie de prolongée spectrum- lactamase productrices
Escherichia coli
ST131 est entraîné par un seul sous-clone hautement pathogène,
H
30-Rx. MBIO 4 (6): e00377-13. doi: 10.1128 / mBio.00377-13.L'épidémie de prolongée spectrum- lactamase productrices
Escherichia coli
ST131 est entraîné par un seul sous-clone hautement pathogène,
H
30-Rx. MBIO 4 (6): e00377-13. doi: 10.1128 / mBio.00377-13.L'épidémie de prolongée spectrum- lactamase productrices
Escherichia coli
ST131 est entraîné par un seul sous-clone hautement pathogène,
H
30-Rx. MBIO 4 (6): e00377-13. doi: 10.1128 / mBio.00377-13.L'épidémie de prolongée spectrum- lactamase productrices
Escherichia coli
ST131 est entraîné par un seul sous-clone hautement pathogène,
H
30-Rx. MBIO 4 (6): e00377-13. doi: 10.1128 / mBio.00377-13.
Éditeur Julian Parkhill, l'Institut SangerÉditeur Julian Parkhill, l'Institut Sanger
droits d'auteur © 2013 Prix et al. Ceci est un article en accès libre distribué sous les termes de la Creative Commons Attribution-ShareAlike 3.0 Unported , Ce qui permet une utilisation non commerciale sans restriction, la droits d'auteur © 2013 Prix et al. Ceci est un article en accès libre distribué sous les termes de la Creative Commons Attribution-ShareAlike 3.0 Unported , Ce qui permet une utilisation non commerciale sans restriction, la droits d'auteur © 2013 Prix et al. Ceci est un article en accès libre distribué sous les termes de la Creative Commons Attribution-ShareAlike 3.0 Unported , Ce qui permet une utilisation non commerciale sans restriction, la droits d'auteur © 2013 Prix et al. Ceci est un article en accès libre distribué sous les termes de la Creative Commons Attribution-ShareAlike 3.0 Unported , Ce qui permet une utilisation non commerciale sans restriction, la
distribution et la reproduction sur tout support, à condition que l'auteur original et la source sont crédités. Adresse de correspondance Lance B. Prix, [email protected].
H le transfert de gènes orizontal est l'une des plus puissantes forcesH le transfert de gènes orizontal est l'une des plus puissantes forces
dans l'évolution bactérienne. Le potentiel de transformation de ce processus est
peut-être mieux fi é exemplifié par l'acquisition des déterminants de la résistance aux
antimicrobiens: dans un événement génétique unique, une bactérie susceptible
antimicrobien peut acquérir une suite complexe de déterminants de la résistance et
deviennent résistantes aux antimicrobiens multiples. Ainsi, le transfert horizontal fréquent
entre les différents
souches peuvent potentiellement conduire la propagation de la résistance aux antibiotiques
dans la population bactérienne, sans aucun changement dans la répartition des souches.
Cependant, lorsque des clones bactériens virulents acquièrent ces éléments, ils peuvent
rapidement émerger au sein de la population grâce à l'expansion clonale et d'acquérir ainsi
une prédominance locale ou même mondiale (1-3). Quantification de la contribution relative de
l'horizontale (transfert de gènes) et verticale (expansion clonale) méca-
ARTICLE DE RECHERCHE
Novembre / Décembre 2013 Volume 4 Numéro 6 e00377-13 ® mbio.asm.org 1® mbio.asm.org 1® mbio.asm.org 1
on February 22, 2020 by guesthttp://mbio.asm.org/Downloaded from

nismes à l'émergence d'agents pathogènes bactériens
multirésistante fourniront des informations importantes
sur l'évolution de ces agents pathogènes et d'informer les
nouvelles stratégies d'intervention.
En 2008, précédemment non reconnu
Escherichia coli
groupe clonal, le type de séquence 131
Escherichia coli
groupe clonal, le type de séquence 131
(de ST131), a été identi fi ées dans neuf pays répartis
sur trois continents (4,
5). Aujourd'hui, ST131 est le pathogène dominant
extraintestinal
E. coli (
ExPEC) souche dans le monde extraintestinal
E. coli (
ExPEC) souche dans le monde extraintestinal
E. coli (
ExPEC) souche dans le monde
entier, mais les analyses rétrospectives suggèrent
que l'émergence d'une pandémie ST131 a eu lieu sur
une période de moins de 10 ans (6, 7). ST131 fait
partie du groupe phylogénique virulent et B2 a été
rapporté pour provoquer un large éventail d'infections,
y compris la méningite, l'ostéomyélite, la myosite,
orchiépididymite et péritonite (6, 8-10). Cependant,
ST131 est le plus souvent associée à une infection
des voies urinaires (UTI) et est un important agent
étiologique des infections urinaires, les infections
rénales et urosepsis aux États-Unis et au niveau
international (11-
14). La génétique des populations analyse des isolats
ST131 a indiqué que la récente propagation épidémique
de ce groupe est entraîné par les descendants d'une
seule souche, nommée sous-clone
H
30, qui ne diffèrent seule souche, nommée sous-clone
H
30, qui ne diffèrent seule souche, nommée sous-clone
H
30, qui ne diffèrent
des membres des autres, ST131 moins répandues par les
sous-clones de transport
fi mH
30, un allèle du gène sous-clones de transport
fi mH
30, un allèle du gène sous-clones de transport
fi mH
30, un allèle du gène
codant pour la mannose-spéci fi ques de type 1 fi mbrial
adhésine, FimH (15).
Au cours de la dernière décennie, l'émergence de
souches multirésistantes ExPEC a fait un traitement
UTI plus problématique, conduisant à une
antibiothérapie discordante et augmentation de la
morbidité et de la mortalité (16-19). Cette augmentation
multirésistante a IVU été en grande partie en raison de
l'augmentation rapide de la prévalence de ExPEC
les souches, en particulier de
déterminants hébergeant ST131 pour
à spectre étendu lactamases (ESBL) et à spectre étendu lactamases (ESBL) et
la résistance à triméthoprime
le sulfaméthoxazole et le fl uoroquinolones (FQ) (16,
20-27).
Le CTX-M-15 lactamase est leLe CTX-M-15 lactamase est le
dominante BLSE dans ST131 et est de plus en plus trouvé
dans les isolats à la fois provoquant une infection urinaire et
la bactériémie (13, 28-31). La phylogénie des gènes CTX-M
suggère que ces enzymes ont été soulevées par la
mobilisation des chromosomes
- lactamase (
bLA)
gènes- lactamase (
bLA)
gènes- lactamase (
bLA)
gènes
de l'organisme commensale intestinale
Kluyvera (
32). Un de l'organisme commensale intestinale
Kluyvera (
32). Un de l'organisme commensale intestinale
Kluyvera (
32). Un
certain nombre d'enzymes CTX-M ont augmenté à la
prévalence depuis les années 1990, avec un nouveau type
CTX-M apparaissant fréquemment dans plusieurs pays
lointains sí-
CD462 JJ2230 JJ2050 JJ2193 JJ2578 JJ2537 JJ2183 QU061 JJ2421 JJ2430 Calg56 (MH19502) JJ2135 JJ2006 MHVlab2 JJ1975 JJ2194 JJ2265 JJ2444-XXXXXXXX JJ2040 JJ2057 MHHDE2 QU189 MH5753 JJ2606 CD338 JJ1915 JJ2206 JJ2216 JJ2196 JJ2210 CD449 CD451 CD460 JJ2142 CU790 JJ2117 JJ2641 CU799 JJ2489-XXXXXXX JJ2120 JJ2539 CD453 JJ2286 CU797 JJ2004 CD257 CD459 JJ1962 JJ2350 JJ2119 JJ2251 JJ1968 JJ1952 CD458 JJ2309 JJ2033 JJ2408 JJ2579 JJ1950 JJ1972 JJ2163 JJ2351 JJ2500 JJ2580 QUC10 JJ2185 JJ2577 JJ2459 JJ2433 CD340 JJ2508 JJ1902 JJ2587 JJ2063 JJ2220 JJ2062 JJ2239 JJ2308 JJ2630 QU153 JJ2631 JJ2125 JJ2152 JJ2064 QU221 QU192 CU758 CU761 JJ2674 JJ2334 JJ2545 JJ2192 QUC01 JJ2642 JJ2317 QU008 JJ2519 JJ1887-XXXXX JJ2667 JJ2218 JJ1933 JJ2247 JJ1867 JJ2603 JJ2130 JJ1886-XXXXXXXX JJ2662 JJ1995 JJ2048 JJ2254 JJ2255 JJ2352 CD454 JJ1967 JJ2534 QU162 JJ2184 CD358-XXXXXXXMH5800-XXXXXXX JJ1974 QUC09 QUC03 QUC04 JJ2047 JJ1900 JJ2044 QUC08 JJ1895 JJ2226 JJ2306 JJ2287 JJ2224 JJ2507 MH15802 QU021 JJ2611 JJ1990 JJ1976 JJ2128 QU048 JJ2225 JJ1934 JJ2312 JJ2495 JJ2195 JJ1877 JJ2269 QU198 JJ2289 JJ2570 JJ2227 JJ2013 JJ2180 CD263 CD400 JJ1942 JJ2540 MH16102 JJ2441-XXXXXX JJ1928 JJ1930 JJ2613 JJ2436 Calg06 (MH1100) JJ2453 JJ2622 JJ2434XXXXX JJ2627 QUC02-XXXXXX CD452 JJ2483 JJ2419 JJ2045 MHMECB5 MHVA1 JJ2333 JJ2544 CD254 JJ1905 JJ2250 PH0872 JJ2032 CD261 JJ1903 JJ2150 JJ2271 JJ2041 JJ1966 JJ2145 JJ2214 CD260 JJ2212 JJ2213 JJ2332 QU161 JJ2176 JJ1977 QU110 JJ1906 JJ2221 JJ2201 JJ2277 QU233 QU206 JJ2178 JJ2190 JJ2015 JJ2001 JJ2136 JJ2362 JJ2553 JJ2285 JJ2291 JJ2191 JJ2122 JJ2321 PH53135 JJ2325 JJ2324 CD252 CD255 JJ1961 JJ2005 JJ2353 JJ2098 JJ2407 JJ2535 CD253 CD256 JJ1904 JJ1894 CD259 JJ2143 Calg51 (MH17102) -XXXXXXXX JJ2624JJ2146 JJ2199 JJ2132 JJ1959 JJ2648 JJ2188 JJ1918 JJ2197 JJ2203 JJ2341 JJ2207 JJ1908 JJ2133 JJ1907 JJ2297 JJ1896 JJ2259 JJ2205 JJ2346 JJ2345 JJ1899 JJ2017 JJ2018 JJ2526 JJ2124 JJ2171 JJ2252 JJ1980 JJ2069 QU249 JJ2425 JJ1914-XXXXXXXX JJ2008-XXXXXXX CMB106 JJ2481 JJ1991 JJ1879 JJ2604 JJ2296 JJ2087 MHHDE1 MHHBS4 MHVB8 MH3756 MH6373 JJ2422 JJ2427 JJ2320 JJ2187 JJ1920 JJ2242 JJ2258 JJ2640 JJ2647 JJ2264 JJ2262 JJ2266 JJ2566 JJ2245 JJ2605 JJ2639 JJ1951 JJ1993 JJ2241 JJ2410 QU073 QU102 QU015 JJ2053 MHFV7569 MHFV7595 CMB107 JJ2007 QU106 JJ2243-XXXXXX MH5936 JJ2546 JJ2548 JJ2547-XXXXXXXX JJ2618 JJ2621 JJ1922 JJ1923 JJ2650 JJ2657-XXXXXXXX JJ2260 JJ2237 JJ2307 JJ2559 JJ2560 JJ2335 JJ2490 JJ2166 JJ2484 JJ2208 CD311 CD345 CD313 CD408 JJ2009 JJ2010 QU240 QU200 JJ2118-XXXXXXXX CD505 JJ1913 JJ2532 JJ2521 JJ2256 JJ1916 JJ2000 MHHDE3 JJ2420 JJ2038-XXXXXXXX CD301MHKN1604-XXXXXXX CD288 CD437 CD437 JJ2055 QUC12 QU019 JJ2514 QU245 JJ2516 CD407 JJ1996 CD413 CD287 JJ2550 CD467 CD559 CD436 JJ1998 CD352 CD434 CD319 CD433 CD471 CD336 CD551 CD412 CD404 CD361 CD461 CD536 CD285 CD303 CD455 CD466 CD417 CD351 PH0168 CD394 CD347 CD456 CD264 JJ1901 CD265 QU259 QU289 QU268 QU270 QU271 QU300 QU252 QU253 QU251 QU255 QU258 CD262 QU299 QUC13 CD306 JJ2244-XXXXXXX CD424 MHAH10 MHAH15 MHAH9 JJ2127 JJ1880 JJ2633 JJ2169 JJ2635 JJ2141 CD297 CD296 CD325 JJ2608 Calg30 (MH8501) JJ2637 JJ2037 JJ1958 JJ2502 JJ2659 JJ2517 CD545 CD548 CD549 CD550 CD375 CD544 CD390 CD512 CD553 CD539 CD541 CD383 CD382 CD356 CD498 JJ1970 JJ1971 JJ1917 JJ2054 JJ2012 JJ2358 JJ2555-XXXXXXXX JJ2417 JJ2575 JJ2615 JJ2003 JJ1910 JJ1911 JJ1912 JJ2223 JJ1878 JJ2424 JJ2426 JJ2449 JJ2428 JJ2429 JJ2442 JJ2437 JJ2450 JJ2574 JJ2413 JJ2416 JJ2232 JJ2649 JJ2431 JJ2432JJ2452 JJ2011 JJ2643-XXXXXXX JJ2645 JJ2646 CMB114 JJ2451 JJ2030 JJ2134 JJ2179 JJ2240 B1-2-1 (H17) JJ2591-XXXXXX JJ2592 QU264 JJ2663 QU090 CD280 CD365 JJ2029 JJ2039 JJ2474 JJ2209 JJ1909 JJ1919 JJ2528 JJ2463 JJ2594 JJ2014 JJ1997 JJ2457 CD268 CD266 CD331 JJ2016 JJ1969 CD357 CD318 JJ1999 JJ2576 CD249 CD250 CD251 JJ2668 JJ1932 JJ1897 JJ2002
300 SNPs
H
30
H
30
900 SNPs
900 SNPs
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
35
35
30
30
30
30
30
30
30
30
22
22
27
22
22
22
22
22
22
22
22
22
35
94
22
22
35
30
22
15
22
22
22
31
22
22
22
22
22
35
22
22
22
22
35
41
41
41
41
41
41
41
UNE. B.
CD467
JJ2555
JJ2444
G132
JJ2041
CD456
JJ2087
JJ2578
JJ2508
MVAST077
QU090
JJ2243
CD340
KN1604
CD471
CU758
JJ2434
CD345
ZH071
CD436
MVAST084
JJ1901
JJ2183
CD306
JJ2591
SaT049
CD390
JJ2134
JJ2118
JJ2038
MVAST131
MVAST0036
JJ1914
SaT158
JJ2193
CD413
MH5800
CD301
QUC12
G150
JJ2050
CD400
MVAST046
JJ2016
JJ1996
H003
CD505
H17
MVAST020
JJ2547
CD466
JJ2009
CD249
H061
JJ2489
CD331
JJ1969
ZH164
JJ2244
U004
JJ1886
SaT142
JJ2608
MVAST179
MVAST038
JJ2528
CD303
G199
SaT040
CD455
ZH193
JJ2643
G216
MVAST158
NA114
QU300
JJ1897
JJ2008
CD311
JMI268
JMI025
H006
MVAST167
JJ2550
ZH063
JJ2657
C001
JJ1887
JJ2007
MH17102
H016
U024
G213
JJ2055
JJ2441
CD347
JJ2210
QUC02
CU799
JJ1999
CD358
CD449
MVAST014
JJ2668
JJ1908
U054
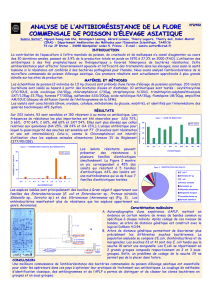
FIG. 1 PFGE dendrogramme et l'ensemble du génome SNP basé phylogénie
E. coli
ST131. (A) à base d'dendrogramme PFGE de
E. coli
isolats FIG. 1 PFGE dendrogramme et l'ensemble du génome SNP basé phylogénie
E. coli
ST131. (A) à base d'dendrogramme PFGE de
E. coli
isolats FIG. 1 PFGE dendrogramme et l'ensemble du génome SNP basé phylogénie
E. coli
ST131. (A) à base d'dendrogramme PFGE de
E. coli
isolats FIG. 1 PFGE dendrogramme et l'ensemble du génome SNP basé phylogénie
E. coli
ST131. (A) à base d'dendrogramme PFGE de
E. coli
isolats FIG. 1 PFGE dendrogramme et l'ensemble du génome SNP basé phylogénie
E. coli
ST131. (A) à base d'dendrogramme PFGE de
E. coli
isolats FIG. 1 PFGE dendrogramme et l'ensemble du génome SNP basé phylogénie
E. coli
ST131. (A) à base d'dendrogramme PFGE de
E. coli
isolats
(ST131
n
(ST131
n
524), tel que déduit à l'intérieur de BioNumerics selon la
Procédé de groupe de paire non pondéré basé sur les coef fi cients de dés. (B) à base de phylogénie SNP-génome entier d'isolats
sélectionnés ST131 (
n
105) et le génome de référence NA114. SNPs ont été identi fi é des régions génomiques équivalant à environ sélectionnés ST131 (
n
105) et le génome de référence NA114. SNPs ont été identi fi é des régions génomiques équivalant à environ sélectionnés ST131 (
n
105) et le génome de référence NA114. SNPs ont été identi fi é des régions génomiques équivalant à environ
44,7% du génome de référence qui a été partagé entre tous les isolats et séquencés à 10 couverture. L'analyse de ces régions 44,7% du génome de référence qui a été partagé entre tous les isolats et séquencés à 10 couverture. L'analyse de ces régions
génomiques partagées révélé
2531-informatif et parcimonie 4000 SNPs au total du génome de base (à l'exclusion des régions acquises à l'horizontale) qui ont
été utilisés pour construire la phylogénie présenté ici. homoplasie indice (HI)
0,012. Le bloc violet met en évidence les
H
30 sous-clone.0,012. Le bloc violet met en évidence les
H
30 sous-clone.0,012. Le bloc violet met en évidence les
H
30 sous-clone.
Prix et al.
2 ® mbio.asm.org ® mbio.asm.org Novembre / Décembre 2013 Volume 4 Numéro 6 e00377-13
FQ-R
CTX-M-15
fimH
complex
fimH
complex
CTX-M-15
FQ-R
70 80
90
100
on February 22, 2020 by guesthttp://mbio.asm.org/Downloaded from

multanément, ce qui suggère des événements de transfert indépendants (33). Ceci, ainsi
que la diversité substantielle des éléments de résistance transférables dans ST131, a
conduit certains à conclure que le transfert horizontal doit être les BLSE de
mechanismwhereby dominants ont pris de l'importance parmi les souches du clone ST131
(7, 34, 35). Comment-
jamais, d'autres preuves suggèrent que l'expansion clonale contribue significativement à la
propagation de la résistance aux antimicrobiens dans les
E. coli
propagation de la résistance aux antimicrobiens dans les
E. coli
(36-39).
Jusqu'à tout récemment, notre connaissance de l'épidémiologie et la dispersion des
souches bactériennes, y compris d'origine ST131, est basée
9 SNPs
H17
G213
JJ2193
NA114 (ref)
MH5800
JJ2210
JJ1886
JJ2550
JJ2434
JJ2643
KN1604
CD358
JJ2244
MVAST131
MVAST0036
MVAST179
JJ2444
JJ1887 U004
JJ2578
MVAST038
JJ1914
CD449
MH17102
ZH193
JJ2041
JJ2528
JJ2508
JJ2118
JMI268
G216
CD340
QUC02
JJ1897
MVAST158
U024
JJ2441
JJ2009
JJ2555
C001
JJ2243
JJ2668
JJ2608
MVAST077
JJ2183
JJ2134
H006
G150
JJ2657
SaT158
H016
H003
MVAST046
JJ2008
JJ2007
CU758
SaT049
MVAST084
JJ2050
ZH164
CD306
MVAST014
JJ2038
JJ1908
JJ2547
JJ2489
CU799
H
30-Rx
H
30-Rx
H
30
H
30
H
30-R
H
30-R
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
30
35
35
30
30
30
30
30
30
30
30
30
30
22
22
27
FIG2 analyse phylogénétique haute résolution de l'émergence de la résistance à la fl uoroquinolone production andCTX-M-15. Environ 51,8% du génome de référence a été partagé entre tous les isolats et FIG2 analyse phylogénétique haute résolution de l'émergence de la résistance à la fl uoroquinolone production andCTX-M-15. Environ 51,8% du génome de référence a été partagé entre tous les isolats et
séquencée à 10 couverture. L'analyse de ces régions génomiques partagées a révélé 72 SNPs informatif et parcimonie 771 SNPs au total lesemployés génome base (hors régions acquises à l'horizontale) qui séquencée à 10 couverture. L'analyse de ces régions génomiques partagées a révélé 72 SNPs informatif et parcimonie 771 SNPs au total lesemployés génome base (hors régions acquises à l'horizontale) qui
ont été utilisés pour construire la phylogénie présenté ici. homoplasie indice (HI)
0.000. Les blocs de couleur mettent en évidence les trois sous-clones ST131 imbriqués,
H
30 (violet),
H
30-R (bleu),
H
30-Rx (jaune).0.000. Les blocs de couleur mettent en évidence les trois sous-clones ST131 imbriqués,
H
30 (violet),
H
30-R (bleu),
H
30-Rx (jaune).0.000. Les blocs de couleur mettent en évidence les trois sous-clones ST131 imbriqués,
H
30 (violet),
H
30-R (bleu),
H
30-Rx (jaune).0.000. Les blocs de couleur mettent en évidence les trois sous-clones ST131 imbriqués,
H
30 (violet),
H
30-R (bleu),
H
30-Rx (jaune).0.000. Les blocs de couleur mettent en évidence les trois sous-clones ST131 imbriqués,
H
30 (violet),
H
30-R (bleu),
H
30-Rx (jaune).0.000. Les blocs de couleur mettent en évidence les trois sous-clones ST131 imbriqués,
H
30 (violet),
H
30-R (bleu),
H
30-Rx (jaune).0.000. Les blocs de couleur mettent en évidence les trois sous-clones ST131 imbriqués,
H
30 (violet),
H
30-R (bleu),
H
30-Rx (jaune).
Clonale extension de céphalosporine résistant
E. coli
Clonale extension de céphalosporine résistant
E. coli
Novembre / Décembre 2013 Volume 4 Numéro 6 e00377-13 ® mbio.asm.org 3® mbio.asm.org 3® mbio.asm.org 3
FQ-R
CTX-M-15
fimH
complex
fimH
complex
on February 22, 2020 by guesthttp://mbio.asm.org/Downloaded from

en grande partie sur le typage de séquence (MLST) multilocus, qui a une résolution
limitée au niveau du sous-clone, et sur l'analyse électrophorèse sur gel puisée fi eld
(PFGE), qui est très vulnérable aux distorsions des événements de transfert de gène
horizontal et l'interprétation subjective. Dans l'étude actuelle, nous avons utilisé
l'analyse du polymorphisme singlenucleotide du génome entier (SNP) pour reconstruire
la phylogénie de ST131 et les déterminants de la résistance superposée et la
sensibilité phénotypique sur cette phylogénie pour élucider l'histoire de l'évolution de la
fl résistance uoroquinolone et la production BLSE dans ce pathogène important.
RÉSULTATS
analyse PFGE. Une collection de 524 isolats ST131 cultivée à partir humains et les analyse PFGE. Une collection de 524 isolats ST131 cultivée à partir humains et les
animaux entre 1967 et 2011 a été analysée par électrophorèse en champ pulsé, ce qui a
abouti à un dendrogramme complexe (Fig. 1A) .dans les dendrogramme basé PFGE, les
isolats qui étaient résistants fl uoroquinolone et / ou
bla
CTX-M-15 positif, bien que isolats qui étaient résistants fl uoroquinolone et / ou
bla
CTX-M-15 positif, bien que isolats qui étaient résistants fl uoroquinolone et / ou
bla
CTX-M-15 positif, bien que isolats qui étaient résistants fl uoroquinolone et / ou
bla
CTX-M-15 positif, bien que
présentant un certain regroupement, ont été entremêlés beaucoup avec ceux qui étaient
fl uoroquinolone sensibles et / ou
bla
CTX-M-15 négatif. A ce titre, l'analyse des rapports fl uoroquinolone sensibles et / ou
bla
CTX-M-15 négatif. A ce titre, l'analyse des rapports fl uoroquinolone sensibles et / ou
bla
CTX-M-15 négatif. A ce titre, l'analyse des rapports fl uoroquinolone sensibles et / ou
bla
CTX-M-15 négatif. A ce titre, l'analyse des rapports
précédents PFGE a soutenu suggérant l'acquisition horizontale fréquente des
déterminants de la résistance fl uoroquinolone et
bla
CTX-M-15 entre les différents déterminants de la résistance fl uoroquinolone et
bla
CTX-M-15 entre les différents déterminants de la résistance fl uoroquinolone et
bla
CTX-M-15 entre les différents déterminants de la résistance fl uoroquinolone et
bla
CTX-M-15 entre les différents
sous-clones de ST131.
reconstruction phylogénétique ST131 base SNP-de-génome entier. De la reconstruction phylogénétique ST131 base SNP-de-génome entier. De la
collection totale qui a subi une analyse de PFGE, 105 ST131 isolats ont été
systématiquement sélectionné pour le séquençage génomique selon des critères fi
ées toprespeci que emphasizeddiversity de fonds génétiques selon la PFGE. Les
105 isolats, qui proviennent de cinq pays et 23 États et provinces au Canada et aux
États-Unis, inclus 22 CTX-M-15 produisant des isolats, qui ont été largement
distribués à travers le dendrogramme électrophorétique (fig. 1A).
comparaisons génomiques identi fi ées SNP loci qui étaient présents dans tous les
isolats et, par conséquent, d'information pour la reconstruction phylogénétique. Le premier
arbre phylogénétique inclus souche non ST131 AA86 (groupB2; ST1876) (40) anoutgroup, à
la racine de l'arbre et pour identifier les clones de base au sein de la phylogénie ST131 (voir
figure S1 dans le matériau supplémentaire.). Ensuite, la souche AA86 a été exclue et une
nouvelle matrice de SNP et arbre phylogénétique ont été générées (voir Fig. S2 dans le
matériau supplémentaire). Depuis AA86 de souche (éloignée) manque certaines des régions
génomiques trouvées dans le clone ST131, l'exclusion de AA86 a augmenté le nombre de
régions génomiques partagées dans l'alignement de séquence et, par conséquent,
augmenté le nombre de SNPs d'information avec qui pour résoudre la phylogénie de ST131.
L'indice de homoplasie (HI) pour ces deux arbres initial (voir Fig. S1 et S2) était
extrêmement élevé ( 0,33), indiquant une recombinaison substantielle. reconstructions extrêmement élevé ( 0,33), indiquant une recombinaison substantielle. reconstructions
phylogénétiques qui incluent des régions génomiques acquises par transfert horizontal
de gènes ne représentent pas avec précision l'histoire de l'évolution des organismes
clonales. Toutefois, ces phylogénie peuvent être utilisés pour identifier les régions
acquises horizontalement. Cela a été accompli ici bymapping au génome de référence
pour les valeurs HI SNPs individuelles, qui ont révélé quatre grandes régions
recombinantes représentant près de 31% du génome.
Exclusion de SNP des quatre régions acquises à l'horizontale conduit à des
arbres withminimal homoplasie (indice de homoplasie [HI]
0,012) (voir Fig. S3 dans le matériau supplémentaire), suggérant
phylogénie de haute précision (41). La figure 1B montre le SNP du génome entier
résultant phylogénie pour les 105 isolats ST131, plus la souche ST131 NA114 génome
de référence (42).
base-génome ensemble regroupement des sous-clones résistants. La phylogénie base-génome ensemble regroupement des sous-clones résistants. La phylogénie
base SNP du génome entier a montré des groupements distincts de souches portant spéci fi
ques
fi mH
alleles (Fig. 1B), ainsi que
gyrA
ques
fi mH
alleles (Fig. 1B), ainsi que
gyrA
ques
fi mH
alleles (Fig. 1B), ainsi que
gyrA
ques
fi mH
alleles (Fig. 1B), ainsi que
gyrA
et
parC
et les allèles de type O (voir l'ensemble de données S1 et S1 dans le tableau et
parC
et les allèles de type O (voir l'ensemble de données S1 et S1 dans le tableau et
parC
et les allèles de type O (voir l'ensemble de données S1 et S1 dans le tableau
matériau supplémentaire). En particulier les souches, portant le
fi mH
30 allele matériau supplémentaire). En particulier les souches, portant le
fi mH
30 allele matériau supplémentaire). En particulier les souches, portant le
fi mH
30 allele
regroupés en un seul clade à faible diversité, désigné
H
30, qui comprend 58 (95%) regroupés en un seul clade à faible diversité, désigné
H
30, qui comprend 58 (95%) regroupés en un seul clade à faible diversité, désigné
H
30, qui comprend 58 (95%)
des 61 isolats de fl uoroquinolone résistant. De plus, presque tous les isolats, malgré
productrices de CTX-M-15 paraissant avoir diverses origines génétiques selon le
dendrogramme basé PFGE (Fig. 1A), regroupées en une subclade distincte au sein de
la
H
30 clade (Fig. 1B).la
H
30 clade (Fig. 1B).la
H
30 clade (Fig. 1B).
Pour résoudre davantage l'histoire de l'évolution de la
H
30 sous-clone, des Pour résoudre davantage l'histoire de l'évolution de la
H
30 sous-clone, des Pour résoudre davantage l'histoire de l'évolution de la
H
30 sous-clone, des
séquences génomiques de la 64
H
30 isolats et leurs trois voisins les plus proches ont séquences génomiques de la 64
H
30 isolats et leurs trois voisins les plus proches ont séquences génomiques de la 64
H
30 isolats et leurs trois voisins les plus proches ont
été analysés séparément du reste des isolats (Fig. 2). L'alignement de ces
séquences au fi nis génome de référence NA114 augmenté le nombre de
sharednucleotides et a révélé SNPs d'information supplémentaires qui ont été utilisés
pour générer la haute résolution et haute précision (HI
0,000) phyloge-
arbre Netic représenté sur la figure. 2. Cet arbre a suggéré que l'acquisition de la
fi mH
30 arbre Netic représenté sur la figure. 2. Cet arbre a suggéré que l'acquisition de la
fi mH
30 arbre Netic représenté sur la figure. 2. Cet arbre a suggéré que l'acquisition de la
fi mH
30
allele précédée de l'acquisition de la résistance à la fl uoroquinolone par un seul ancêtre
au sein de la
H
30 sous-clone, qui a été suivie par une grande expansion clonale de fl au sein de la
H
30 sous-clone, qui a été suivie par une grande expansion clonale de fl au sein de la
H
30 sous-clone, qui a été suivie par une grande expansion clonale de fl
uoroquinolone résistant
H
30 souches. Pour distinguer les uoroquinoloneresistant fl liés clonale
H
30
H
30 souches. Pour distinguer les uoroquinoloneresistant fl liés clonale
H
30
H
30 souches. Pour distinguer les uoroquinoloneresistant fl liés clonale
H
30
H
30 souches. Pour distinguer les uoroquinoloneresistant fl liés clonale
H
30
isolats du uoroquinolonesusceptible fl ancestral
H
30 isolats, le sous-clone isolats du uoroquinolonesusceptible fl ancestral
H
30 isolats, le sous-clone isolats du uoroquinolonesusceptible fl ancestral
H
30 isolats, le sous-clone
résistant à l'intérieur
H
30 a été désigné
H
30-R.résistant à l'intérieur
H
30 a été désigné
H
30-R.résistant à l'intérieur
H
30 a été désigné
H
30-R.résistant à l'intérieur
H
30 a été désigné
H
30-R.résistant à l'intérieur
H
30 a été désigné
H
30-R.
Dans ce phylogénie haute résolution, 20 (91%) des 22 isolats qui ST131
effectuée
bla
CTX-M-15 isolats -Incluant de l'Australie, la Corée du Sud, le Portugal, le effectuée
bla
CTX-M-15 isolats -Incluant de l'Australie, la Corée du Sud, le Portugal, le effectuée
bla
CTX-M-15 isolats -Incluant de l'Australie, la Corée du Sud, le Portugal, le effectuée
bla
CTX-M-15 isolats -Incluant de l'Australie, la Corée du Sud, le Portugal, le
Canada et les États-Unis- ont formé une distincte, sous-clone unique ancêtre
dans
H
30-R. En raison de ses caractéristiques de résistance plus étendues, ce
bla
dans
H
30-R. En raison de ses caractéristiques de résistance plus étendues, ce
bla
dans
H
30-R. En raison de ses caractéristiques de résistance plus étendues, ce
bla
dans
H
30-R. En raison de ses caractéristiques de résistance plus étendues, ce
bla
CTXM-15- a été désigné sous-clone associé
H
30-Rx (Fig. 2). Trois SNPs canoniques CTXM-15- a été désigné sous-clone associé
H
30-Rx (Fig. 2). Trois SNPs canoniques CTXM-15- a été désigné sous-clone associé
H
30-Rx (Fig. 2). Trois SNPs canoniques CTXM-15- a été désigné sous-clone associé
H
30-Rx (Fig. 2). Trois SNPs canoniques
distingués
H
30-Rx lesemployés reste de
H
30 rAvec 100% fi délité.distingués
H
30-Rx lesemployés reste de
H
30 rAvec 100% fi délité.distingués
H
30-Rx lesemployés reste de
H
30 rAvec 100% fi délité.distingués
H
30-Rx lesemployés reste de
H
30 rAvec 100% fi délité.distingués
H
30-Rx lesemployés reste de
H
30 rAvec 100% fi délité.
localisation génomique de l'élément CTX-M-15. Plusieurs études antérieures ont localisation génomique de l'élément CTX-M-15. Plusieurs études antérieures ont
rapporté que
bla
CTX-M-15 est positionné sur un plasmide de type IncFII conjugatif dans le rapporté que
bla
CTX-M-15 est positionné sur un plasmide de type IncFII conjugatif dans le rapporté que
bla
CTX-M-15 est positionné sur un plasmide de type IncFII conjugatif dans le rapporté que
bla
CTX-M-15 est positionné sur un plasmide de type IncFII conjugatif dans le
cadre d'un Tn
3-
comme
ISEcp1-Bla
CTXM-15- orf477 élément mobile. Ici, nous avons cadre d'un Tn
3-
comme
ISEcp1-Bla
CTXM-15- orf477 élément mobile. Ici, nous avons cadre d'un Tn
3-
comme
ISEcp1-Bla
CTXM-15- orf477 élément mobile. Ici, nous avons cadre d'un Tn
3-
comme
ISEcp1-Bla
CTXM-15- orf477 élément mobile. Ici, nous avons cadre d'un Tn
3-
comme
ISEcp1-Bla
CTXM-15- orf477 élément mobile. Ici, nous avons cadre d'un Tn
3-
comme
ISEcp1-Bla
CTXM-15- orf477 élément mobile. Ici, nous avons
effectué une
in silico
effectué une
in silico
analyse pour caractériser la structure et la localisation génomique de l'organe mobile
CTX-M-15 parmi les 22
bla
CTX-M-15 isolats positifs ST131. Dans chaque cas,
bla
CTX-M-15 faisait CTX-M-15 parmi les 22
bla
CTX-M-15 isolats positifs ST131. Dans chaque cas,
bla
CTX-M-15 faisait CTX-M-15 parmi les 22
bla
CTX-M-15 isolats positifs ST131. Dans chaque cas,
bla
CTX-M-15 faisait CTX-M-15 parmi les 22
bla
CTX-M-15 isolats positifs ST131. Dans chaque cas,
bla
CTX-M-15 faisait CTX-M-15 parmi les 22
bla
CTX-M-15 isolats positifs ST131. Dans chaque cas,
bla
CTX-M-15 faisait CTX-M-15 parmi les 22
bla
CTX-M-15 isolats positifs ST131. Dans chaque cas,
bla
CTX-M-15 faisait CTX-M-15 parmi les 22
bla
CTX-M-15 isolats positifs ST131. Dans chaque cas,
bla
CTX-M-15 faisait
partie d'un Tn typique
3-
comme
ISEcp1-Bla
CTX-M-15 orf477 élément transposable. Bien partie d'un Tn typique
3-
comme
ISEcp1-Bla
CTX-M-15 orf477 élément transposable. Bien partie d'un Tn typique
3-
comme
ISEcp1-Bla
CTX-M-15 orf477 élément transposable. Bien partie d'un Tn typique
3-
comme
ISEcp1-Bla
CTX-M-15 orf477 élément transposable. Bien partie d'un Tn typique
3-
comme
ISEcp1-Bla
CTX-M-15 orf477 élément transposable. Bien partie d'un Tn typique
3-
comme
ISEcp1-Bla
CTX-M-15 orf477 élément transposable. Bien
que fi no SNPswere ed entre les différents éléments CTX-M-15, les régions fl Anking
bla
CTX-M-15 que fi no SNPswere ed entre les différents éléments CTX-M-15, les régions fl Anking
bla
CTX-M-15 que fi no SNPswere ed entre les différents éléments CTX-M-15, les régions fl Anking
bla
CTX-M-15
ont souvent été dégradés par l'insertion / suppression (non représenté). Les séquences
Illumina court lues sont suf fi pour identifier de manière fiable le site d'insertion de
l'élément pour tous, mais trois des 22 isolats CTX-M-15-positives séquences dans la
présente étude (tableau 1). Le site d'insertion varie parmi les isolats: 13 effectue une
copie unique sur un plasmide de type de IncFII, quatre réalisée d'une seule copie dans
le chromosome, et deux effectué une copie sur le chromosome et une autre copie sur
un plasmide de type IncFII (tableau 1) . En outre, parmi les souches avec un élément
situé dans le chromosome, cinq sites d'insertion chromosomiques distincts ont été
identifiés ed; seulement deux souches, JJ1886 et JJ1887, qui sont les voisins les plus
proches dans la phylogénie SNP (Fig. 2), l'élément effectuées dans le même
emplacement chromosomique (tableau 1).
Association de la production et BLSE
bla
CTX-M-15 avec leAssociation de la production et BLSE
bla
CTX-M-15 avec leAssociation de la production et BLSE
bla
CTX-M-15 avec leAssociation de la production et BLSE
bla
CTX-M-15 avec le
H
30-Rx ST131 sous-clone. Parce que la plupart des isolats dans les arbres
H
30-Rx ST131 sous-clone. Parce que la plupart des isolats dans les arbres
H
30-Rx ST131 sous-clone. Parce que la plupart des isolats dans les arbres
phylogénétiques étaient d'origine historique (c.-à-pré-2009), nous avons évalué la
généralisabilité de l'association observée de la
H
30-Rx sousgénéralisabilité de l'association observée de la
H
30-Rx sousgénéralisabilité de l'association observée de la
H
30-Rx sous
Prix et al.
4 ® mbio.asm.org ® mbio.asm.org Novembre / Décembre 2013 Volume 4 Numéro 6 e00377-13
on February 22, 2020 by guesthttp://mbio.asm.org/Downloaded from

clone avec la production et BLSE
bla
CTX-M-15 en analysant les isolats cliniques plus clone avec la production et BLSE
bla
CTX-M-15 en analysant les isolats cliniques plus clone avec la production et BLSE
bla
CTX-M-15 en analysant les isolats cliniques plus clone avec la production et BLSE
bla
CTX-M-15 en analysant les isolats cliniques plus
récents, à savoir de 2011 à 2013. Pour ce faire, un total de 261 isolats ST131 de
Seattle, WA, Minneapolis, MN, et Münster, en Allemagne, ont été évalués pour
fi mH
Type Seattle, WA, Minneapolis, MN, et Münster, en Allemagne, ont été évalués pour
fi mH
Type Seattle, WA, Minneapolis, MN, et Münster, en Allemagne, ont été évalués pour
fi mH
Type
allèle,
H
30-Rx appartenance à un sous-clone, la production BLSE, et la possession deallèle,
H
30-Rx appartenance à un sous-clone, la production BLSE, et la possession deallèle,
H
30-Rx appartenance à un sous-clone, la production BLSE, et la possession de
bla
CTX-M-15 ( Tableau 2).
bla
CTX-M-15 ( Tableau 2).
bla
CTX-M-15 ( Tableau 2).
Parmi les 261 isolats récents ST131, 174 (67%) appartenaient à la
H
30 Parmi les 261 isolats récents ST131, 174 (67%) appartenaient à la
H
30 Parmi les 261 isolats récents ST131, 174 (67%) appartenaient à la
H
30
sous-clone, alors que le 87 reste (33%) porté l'un de plusieurs autres associés
ST131-
fi mH
allèles, comme décrit récemment (15). Parmi les 174
H
30 isolats, 163 ST131-
fi mH
allèles, comme décrit récemment (15). Parmi les 174
H
30 isolats, 163 ST131-
fi mH
allèles, comme décrit récemment (15). Parmi les 174
H
30 isolats, 163 ST131-
fi mH
allèles, comme décrit récemment (15). Parmi les 174
H
30 isolats, 163 ST131-
fi mH
allèles, comme décrit récemment (15). Parmi les 174
H
30 isolats, 163
(94%) qui étaient fl uoroquinolone résistantes ont été défini comme
H
30-R. (94%) qui étaient fl uoroquinolone résistantes ont été défini comme
H
30-R. (94%) qui étaient fl uoroquinolone résistantes ont été défini comme
H
30-R.
détection de
H
30Rx spéci fi que SNPs a montré
H
30-Rx composé 44 (27%) 163détection de
H
30Rx spéci fi que SNPs a montré
H
30-Rx composé 44 (27%) 163détection de
H
30Rx spéci fi que SNPs a montré
H
30-Rx composé 44 (27%) 163détection de
H
30Rx spéci fi que SNPs a montré
H
30-Rx composé 44 (27%) 163détection de
H
30Rx spéci fi que SNPs a montré
H
30-Rx composé 44 (27%) 163
H
souches 30-R (Tableau 2).
H
souches 30-R (Tableau 2).
Parmi les 44
H
30-Rx isolats, 34 (77%) étaient BLSE produire, et 33 d'entre Parmi les 44
H
30-Rx isolats, 34 (77%) étaient BLSE produire, et 33 d'entre Parmi les 44
H
30-Rx isolats, 34 (77%) étaient BLSE produire, et 33 d'entre
eux réalisés
bla
CTX-M-15. Cela a été très similaire àeux réalisés
bla
CTX-M-15. Cela a été très similaire àeux réalisés
bla
CTX-M-15. Cela a été très similaire àeux réalisés
bla
CTX-M-15. Cela a été très similaire à
le chariot 74% du
bla
CTX-M-15 observée parmi les historiques genomesequenced
H
le chariot 74% du
bla
CTX-M-15 observée parmi les historiques genomesequenced
H
le chariot 74% du
bla
CTX-M-15 observée parmi les historiques genomesequenced
H
le chariot 74% du
bla
CTX-M-15 observée parmi les historiques genomesequenced
H
le chariot 74% du
bla
CTX-M-15 observée parmi les historiques genomesequenced
H
isole 30-Rx (Fig. 2), mais significativement plus élevée que la faible prévalence
de production soit BLSE ou
bla
CTX-M-15 transport observé chez les non-récente
H
30 isolats ST131 (3%) pour chaque
bla
CTX-M-15 transport observé chez les non-récente
H
30 isolats ST131 (3%) pour chaque
bla
CTX-M-15 transport observé chez les non-récente
H
30 isolats ST131 (3%) pour chaque
bla
CTX-M-15 transport observé chez les non-récente
H
30 isolats ST131 (3%) pour chaque
bla
CTX-M-15 transport observé chez les non-récente
H
30 isolats ST131 (3%) pour chaque
caractère,
H
30 mais pas
H
isolats 30-R (9%) pour chaque caractère, et
H
30-R, mais pas
H
caractère,
H
30 mais pas
H
isolats 30-R (9%) pour chaque caractère, et
H
30-R, mais pas
H
caractère,
H
30 mais pas
H
isolats 30-R (9%) pour chaque caractère, et
H
30-R, mais pas
H
caractère,
H
30 mais pas
H
isolats 30-R (9%) pour chaque caractère, et
H
30-R, mais pas
H
caractère,
H
30 mais pas
H
isolats 30-R (9%) pour chaque caractère, et
H
30-R, mais pas
H
caractère,
H
30 mais pas
H
isolats 30-R (9%) pour chaque caractère, et
H
30-R, mais pas
H
caractère,
H
30 mais pas
H
isolats 30-R (9%) pour chaque caractère, et
H
30-R, mais pas
H
caractère,
H
30 mais pas
H
isolats 30-R (9%) pour chaque caractère, et
H
30-R, mais pas
H
30 isolats-Rx (6% et 2% pour les deux traits, respectivement) (tableau 2). Donc,
bla
CTX-M-15 30 isolats-Rx (6% et 2% pour les deux traits, respectivement) (tableau 2). Donc,
bla
CTX-M-15 30 isolats-Rx (6% et 2% pour les deux traits, respectivement) (tableau 2). Donc,
bla
CTX-M-15
ont représenté presque tous les isolats productrices de BLSE dans le
H
30 ont représenté presque tous les isolats productrices de BLSE dans le
H
30 ont représenté presque tous les isolats productrices de BLSE dans le
H
30
Rx-sous-clone; À l'inverse, dans l'ensemble ST131, la
H
30-Rx a représenté la Rx-sous-clone; À l'inverse, dans l'ensemble ST131, la
H
30-Rx a représenté la Rx-sous-clone; À l'inverse, dans l'ensemble ST131, la
H
30-Rx a représenté la
sous-clone grande majorité de la production BLSE et, en particulier,
bla
CTX-M-15 le chariot. De plus, cette association étroite entre
bla
CTX-M-15 le chariot. De plus, cette association étroite entre
bla
CTX-M-15 le chariot. De plus, cette association étroite entre
H
30-Rx et
bla
CTX-M-15 se vérifiait dans les différents laboratoires qui ont fourni les récents
H
30-Rx et
bla
CTX-M-15 se vérifiait dans les différents laboratoires qui ont fourni les récents
H
30-Rx et
bla
CTX-M-15 se vérifiait dans les différents laboratoires qui ont fourni les récents
H
30-Rx et
bla
CTX-M-15 se vérifiait dans les différents laboratoires qui ont fourni les récents
H
30-Rx et
bla
CTX-M-15 se vérifiait dans les différents laboratoires qui ont fourni les récents
isolats cliniques (données non présentées).
prévalence démographique, géographique et clinique
H
30Rx. Nous avons prévalence démographique, géographique et clinique
H
30Rx. Nous avons prévalence démographique, géographique et clinique
H
30Rx. Nous avons prévalence démographique, géographique et clinique
H
30Rx. Nous avons
également évalué la prévalence relative des
H
30-R etégalement évalué la prévalence relative des
H
30-R etégalement évalué la prévalence relative des
H
30-R et
H
30-Rx sous-clones au sein de la population totale ST131 par rapport à la population
H
30-Rx sous-clones au sein de la population totale ST131 par rapport à la population
des patients et locale en comparant les isolats d'urine de GroupHealth Cooperative à
Seattle, WA, qui dessert une population ambulatoire presque exclusivement, avec de
l'urine isole les laboratoires fromhospital dans theUnited États andGermany que
servemixed patients hospitalisés et externes populations. La prévalence relative des
H
30-Rx était le plus élevé parmi les isolats des hôpitaux allemands (où il a dépassé la
H
30-Rx était le plus élevé parmi les isolats des hôpitaux allemands (où il a dépassé la
prévalence même d'autres
H
isolats 30-R), intermédiaire entre les isolats hospitaliers basés prévalence même d'autres
H
isolats 30-R), intermédiaire entre les isolats hospitaliers basés prévalence même d'autres
H
isolats 30-R), intermédiaire entre les isolats hospitaliers basés
aux États-Unis, et le plus bas parmi les isolats de consultation externe en santé du Groupe
(tableau 3).
Les données relatives à la présence / absence de septicémie cliniquement diagnostiqué
étaient disponibles pour 162 des récentes États-Unis ST131 isolats cliniques, parmi lesquels 12
patients source (7%) Dans l'ensemble ont été diagnostiqués avec une septicémie (tableau 4),
une valeur similaire à la prévalence globale de 5,2% de septicémie diagnostiqué chez les 1.133
isolats cliniques extraintestinales fromwhich les 162 ST131 souches sont dérivées (
P
isolats cliniques extraintestinales fromwhich les 162 ST131 souches sont dérivées (
P
0,26) (15). Cependant, la septicémie a été diagnostiquée chez 28% des PA-
tients avec un
H
30-Rx isolât (tableau 4), une significativement plus grande proportion tients avec un
H
30-Rx isolât (tableau 4), une significativement plus grande proportion tients avec un
H
30-Rx isolât (tableau 4), une significativement plus grande proportion
que chez les patients atteints d'un non-
H
30-Rx,
H
isolât 30-R (6%;
P
0,02), un non-
H
30, que chez les patients atteints d'un non-
H
30-Rx,
H
isolât 30-R (6%;
P
0,02), un non-
H
30, que chez les patients atteints d'un non-
H
30-Rx,
H
isolât 30-R (6%;
P
0,02), un non-
H
30, que chez les patients atteints d'un non-
H
30-Rx,
H
isolât 30-R (6%;
P
0,02), un non-
H
30, que chez les patients atteints d'un non-
H
30-Rx,
H
isolât 30-R (6%;
P
0,02), un non-
H
30, que chez les patients atteints d'un non-
H
30-Rx,
H
isolât 30-R (6%;
P
0,02), un non-
H
30, que chez les patients atteints d'un non-
H
30-Rx,
H
isolât 30-R (6%;
P
0,02), un non-
H
30, que chez les patients atteints d'un non-
H
30-Rx,
H
isolât 30-R (6%;
P
0,02), un non-
H
30, que chez les patients atteints d'un non-
H
30-Rx,
H
isolât 30-R (6%;
P
0,02), un non-
H
30,
ST131 isolât (4%;
P
0,01), toute non
H
30-Rx, ST131 isolât (5%;
P
0,005), ou un isolât ST131 isolât (4%;
P
0,01), toute non
H
30-Rx, ST131 isolât (5%;
P
0,005), ou un isolât ST131 isolât (4%;
P
0,01), toute non
H
30-Rx, ST131 isolât (5%;
P
0,005), ou un isolât ST131 isolât (4%;
P
0,01), toute non
H
30-Rx, ST131 isolât (5%;
P
0,005), ou un isolât ST131 isolât (4%;
P
0,01), toute non
H
30-Rx, ST131 isolât (5%;
P
0,005), ou un isolât ST131 isolât (4%;
P
0,01), toute non
H
30-Rx, ST131 isolât (5%;
P
0,005), ou un isolât ST131 isolât (4%;
P
0,01), toute non
H
30-Rx, ST131 isolât (5%;
P
0,005), ou un isolât
non-ST131 (5,6%;
P
0,003). Pour
H
30 isolats-Rx par rapport à d'autres isolats de ST131, non-ST131 (5,6%;
P
0,003). Pour
H
30 isolats-Rx par rapport à d'autres isolats de ST131, non-ST131 (5,6%;
P
0,003). Pour
H
30 isolats-Rx par rapport à d'autres isolats de ST131, non-ST131 (5,6%;
P
0,003). Pour
H
30 isolats-Rx par rapport à d'autres isolats de ST131, non-ST131 (5,6%;
P
0,003). Pour
H
30 isolats-Rx par rapport à d'autres isolats de ST131,
le risque relatif de la septicémie associée était de 7,5 (95% con fi ance intervalle, de 2,3
à 23,8).
DISCUSSION
Les résultats de cette étude fournissent des preuves convaincantes que l'expansion
clonale est le mécanisme dominant pour la prolifération des
TABLEAU 1 CTX-M-15 emplacements d'éléments parmi les 22
Escherichia coli
TABLEAU 1 CTX-M-15 emplacements d'éléments parmi les 22
Escherichia coli
TABLEAU 1 CTX-M-15 emplacements d'éléments parmi les 22
Escherichia coli
isole ST131 nom
Isoler
Le sous-clone emplacement génomique
une
emplacement génomique
une
site d'insertion
chromosomal
b
chromosomal
b
JJ2444
H
30-Rx
H
30-Rx Chromosome 2037134
JJ2038
H
30-Rx
H
30-Rx Chromosome 2127735
JJ1886
H
30-Rx
H
30-Rx Chromosome 1473842
JJ1887
H
30-Rx
H
30-Rx Chromosome 1473842
JJ2434
H
30-Rx
H
30-Rx Plasmide et le chromosome 4493369
MH5800
H
30-Rx MH5800
H
30-Rx MH5800
H
30-Rx Plasmide et le chromosome 4191808
JJ2547
H
30-Rx
H
30-Rx Plasmide N / A
JJ2555
H
30-Rx
H
30-Rx Plasmide N / A
JJ2008
H
30-Rx
H
30-Rx Plasmide N / A
JJ1914
H
30-Rx
H
30-Rx Plasmide N / A
JJ2441
H
30-Rx
H
30-Rx Plasmide N / A
JJ2489
H
30-Rx
H
30-Rx Plasmide N / A
JJ2657
H
30-Rx
H
30-Rx Plasmide N / A
JJ2643
H
30-Rx
H
30-Rx Plasmide N / A
U004
H
30-Rx
H
30-Rx Plasmide N / A
CD358
H
30-Rx
H
30-Rx Plasmide N / A
NA1114
H
30-Rx NA1114
H
30-Rx NA1114
H
30-Rx Plasmide
c
Plasmide
c
N / A
MH17102
H
30-Rx MH17102
H
30-Rx MH17102
H
30-Rx Indéterminé N / A
QUC02
H
30-Rx
H
30-Rx Indéterminé N / A
KN1604
H
30-Rx KN1604
H
30-Rx KN1604
H
30-Rx Indéterminé N / A
JJ2244
H
30, non
H
30-Rx plasmide
H
30, non
H
30-Rx plasmide
H
30, non
H
30-Rx plasmide
H
30, non
H
30-Rx plasmide N / A
JJ2591 Non-
H
30 Non-
H
30 Non-
H
30 Plasmide N / A
une
Marqué comme indéterminé si le
in silico
analyse des résultats ambigus.
une
Marqué comme indéterminé si le
in silico
analyse des résultats ambigus.
une
Marqué comme indéterminé si le
in silico
analyse des résultats ambigus.
une
Marqué comme indéterminé si le
in silico
analyse des résultats ambigus.
b
emplacement chromosomal basé sur le génome fermé de JJ1886. NA, non applicable.
b
emplacement chromosomal basé sur le génome fermé de JJ1886. NA, non applicable.
c
Rapporté précédemment.
c
Rapporté précédemment.
TABLEAU 2 Association des ST131 sous-clones présentant des caractéristiques de résistance parmi 261 isolats cliniques récents de
Escherichia coli
ST131 des États-Unis et en AllemagneTABLEAU 2 Association des ST131 sous-clones présentant des caractéristiques de résistance parmi 261 isolats cliniques récents de
Escherichia coli
ST131 des États-Unis et en AllemagneTABLEAU 2 Association des ST131 sous-clones présentant des caractéristiques de résistance parmi 261 isolats cliniques récents de
Escherichia coli
ST131 des États-Unis et en AllemagneTABLEAU 2 Association des ST131 sous-clones présentant des caractéristiques de résistance parmi 261 isolats cliniques récents de
Escherichia coli
ST131 des États-Unis et en Allemagne
trait résistance
La prévalence du trait de résistance, non. (%)
souches total (ST131
n
261)souches total (ST131
n
261)souches total (ST131
n
261)
ST131 sous-clone (s)
Non-
H
30 (
n
87)Non-
H
30 (
n
87)Non-
H
30 (
n
87)Non-
H
30 (
n
87)Non-
H
30 (
n
87)
H
30 (
n
174)
H
30 (
n
174)
H
30 (
n
174)
H
30 (
n
174)
Non-
H
30-R (
n
11)Non-
H
30-R (
n
11)Non-
H
30-R (
n
11)Non-
H
30-R (
n
11)Non-
H
30-R (
n
11)
H
30-R (
n
165) Non-
H
30-Rx
H
30-R (
n
165) Non-
H
30-Rx
H
30-R (
n
165) Non-
H
30-Rx
H
30-R (
n
165) Non-
H
30-Rx
H
30-R (
n
165) Non-
H
30-Rx
H
30-R (
n
165) Non-
H
30-Rx
(
n
119)(
n
119)(
n
119)
H
30-Rx (
n
44)
H
30-Rx (
n
44)
H
30-Rx (
n
44)
H
30-Rx (
n
44)
FQ résistant 163 (62) 0 (0) 0 (0) 119 (100) 44 (100)
une
44 (100)
une
BLSE 45 (17) 3 (3) 1 (9) 7 (6) 34 (77)
b
34 (77)
b
bla
CTX-M-15
bla
CTX-M-15 39 (15) 3 (7) 1 (9) 2 (2) 33 (75)
c
33 (75)
c
une
Pour la fl uoroquinolone (FQ) fraction résistante,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (test exact de Fisher [FET]).
une
Pour la fl uoroquinolone (FQ) fraction résistante,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (test exact de Fisher [FET]).
une
Pour la fl uoroquinolone (FQ) fraction résistante,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (test exact de Fisher [FET]).
une
Pour la fl uoroquinolone (FQ) fraction résistante,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (test exact de Fisher [FET]).
une
Pour la fl uoroquinolone (FQ) fraction résistante,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (test exact de Fisher [FET]).
une
Pour la fl uoroquinolone (FQ) fraction résistante,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (test exact de Fisher [FET]).
b
Pour la prévalence du spectre étendu lactamases (ESBL) production,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
b
Pour la prévalence du spectre étendu lactamases (ESBL) production,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
b
Pour la prévalence du spectre étendu lactamases (ESBL) production,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
b
Pour la prévalence du spectre étendu lactamases (ESBL) production,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
b
Pour la prévalence du spectre étendu lactamases (ESBL) production,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
b
Pour la prévalence du spectre étendu lactamases (ESBL) production,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
b
Pour la prévalence du spectre étendu lactamases (ESBL) production,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
c
Pour la prévalence de
bla
CTX-M-15,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
c
Pour la prévalence de
bla
CTX-M-15,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
c
Pour la prévalence de
bla
CTX-M-15,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
c
Pour la prévalence de
bla
CTX-M-15,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
c
Pour la prévalence de
bla
CTX-M-15,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
c
Pour la prévalence de
bla
CTX-M-15,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
c
Pour la prévalence de
bla
CTX-M-15,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
c
Pour la prévalence de
bla
CTX-M-15,
H
30-Rx par rapport à d'autres isolats de ST131,
P
0,001 (FET).
Clonale extension de céphalosporine résistant
E. coli
Clonale extension de céphalosporine résistant
E. coli
Novembre / Décembre 2013 Volume 4 Numéro 6 e00377-13 ® mbio.asm.org 5® mbio.asm.org 5® mbio.asm.org 5
on February 22, 2020 by guesthttp://mbio.asm.org/Downloaded from
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%