1
Physiologie nerveuse
Chapitre I : Physiologie du neurone
Introduction :
Le système nerveux est un réseau de communication dans l’organisme
qui a trois grands rôles : recevoir, traiter ou intégrer des informations et
envoyer des commandes aux effecteurs. Il constitue un système de contrôle et
d’intégration de l’organisme caractérisé par la rapidité de ces réactions et la
localisation de ces sites d’actions. Il est compose de deux types de cellules de
même origine embryologique étroitement associées : les cellules gliales
(névroglie) et les neurones.
Les cellules gliales :
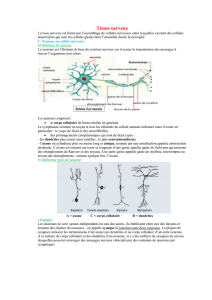
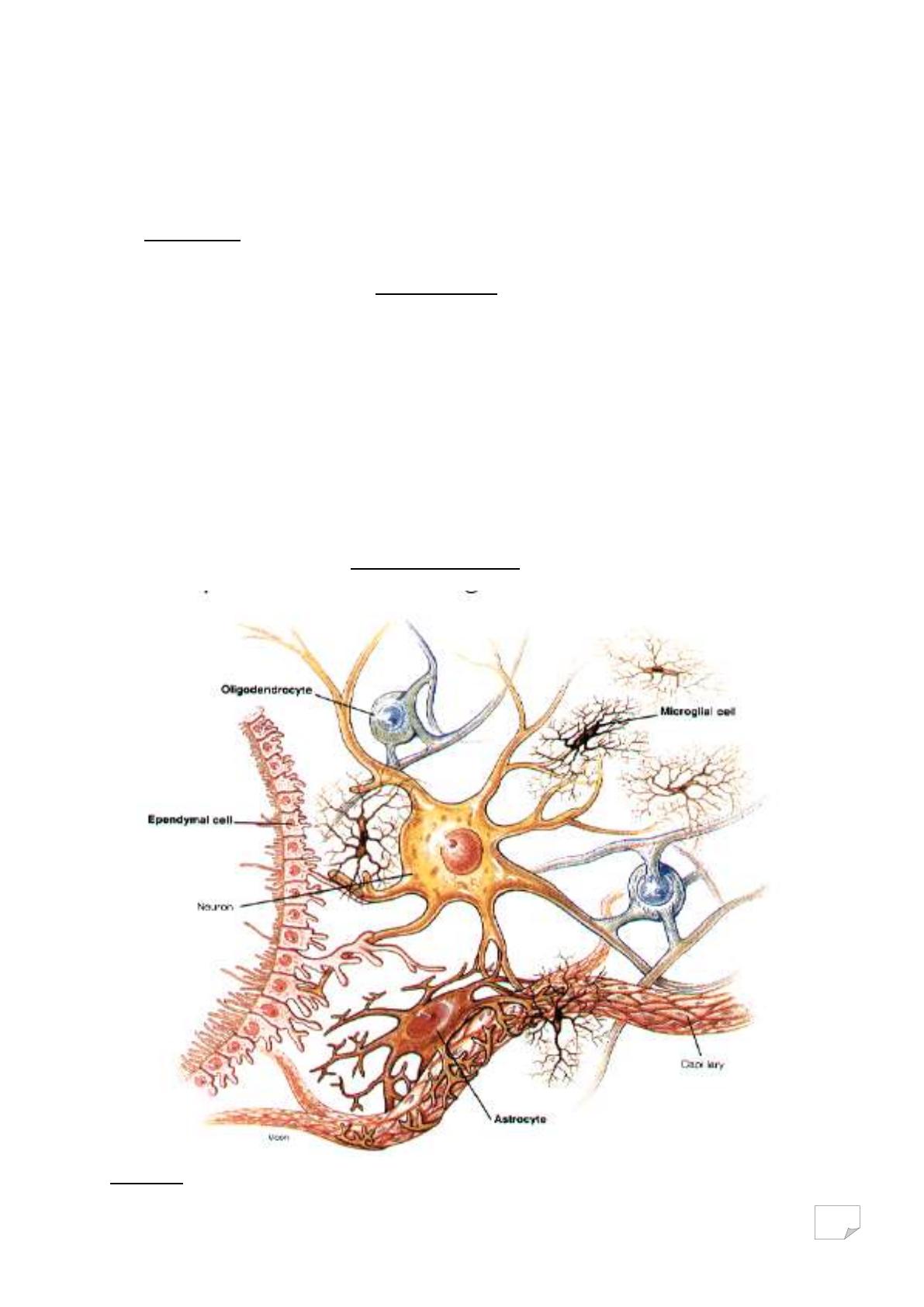
Figure 2 : Vue d’ensemble d’un neurone entouré par les cellules gliales.
2
Ce sont des cellules non excitables qui entourent et protègent les
neurones. Elles sont 10 fois plus nombreuses que les neurones, assurent un
support métabolique par régulation des concentrations ioniques et du pH
extracellulaire, mais aussi, recyclent et synthétisent des précurseurs des
neuromédiateurs. Ce sont des cellules renouvelables qui jouent un rôle de
soutien et de développement du SNC. Il existe cinq types de cellules gliales: les
astrocytes, les microgliocytes, les épendimocytes, les oligodendrocytes(SNC) et
les cellules de Schwann(SNP).
Figure 2 : Vue d’ensemble d’un neurone entouré par les cellules gliales.
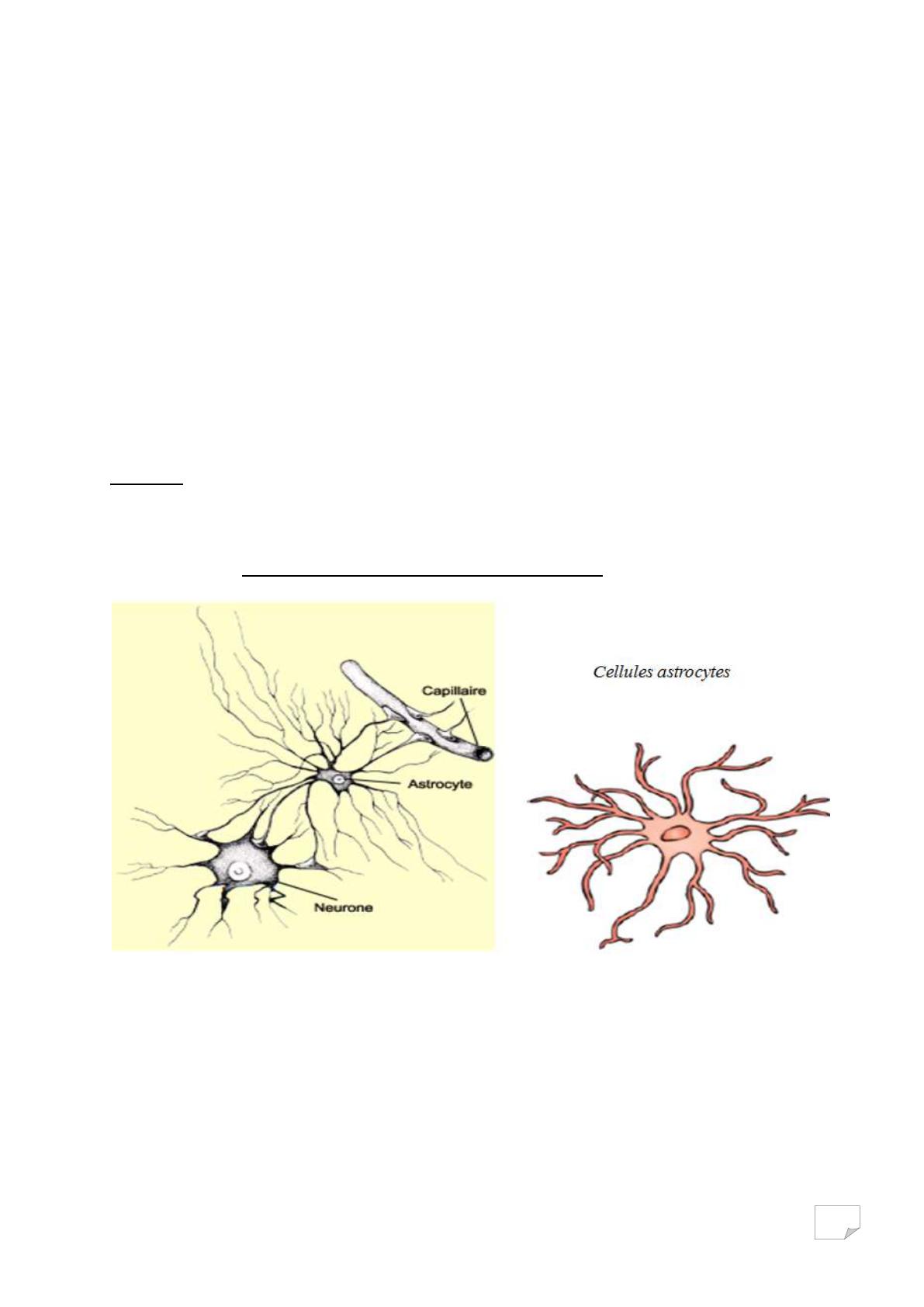
Les astrocytes ou névroglie astrocytaire :
Elles ont un rôle de nutrition et possèdent de nombreuses dendrites qui
partent dans tous les sens (cellules étoilées). Elles envoient des prolongements
vers les vaisseaux sanguins afin de permettre des échanges (nutriments, gaz …)
entre le sang et les neurones. Elles capturent et libèrent les
neurotransmetteurs et sont des éléments constitutifs de la barrière hémato-
encéphalique (isolement des neurones par rapport au sang) et stock le sucre
sous forme de glycogène.

3
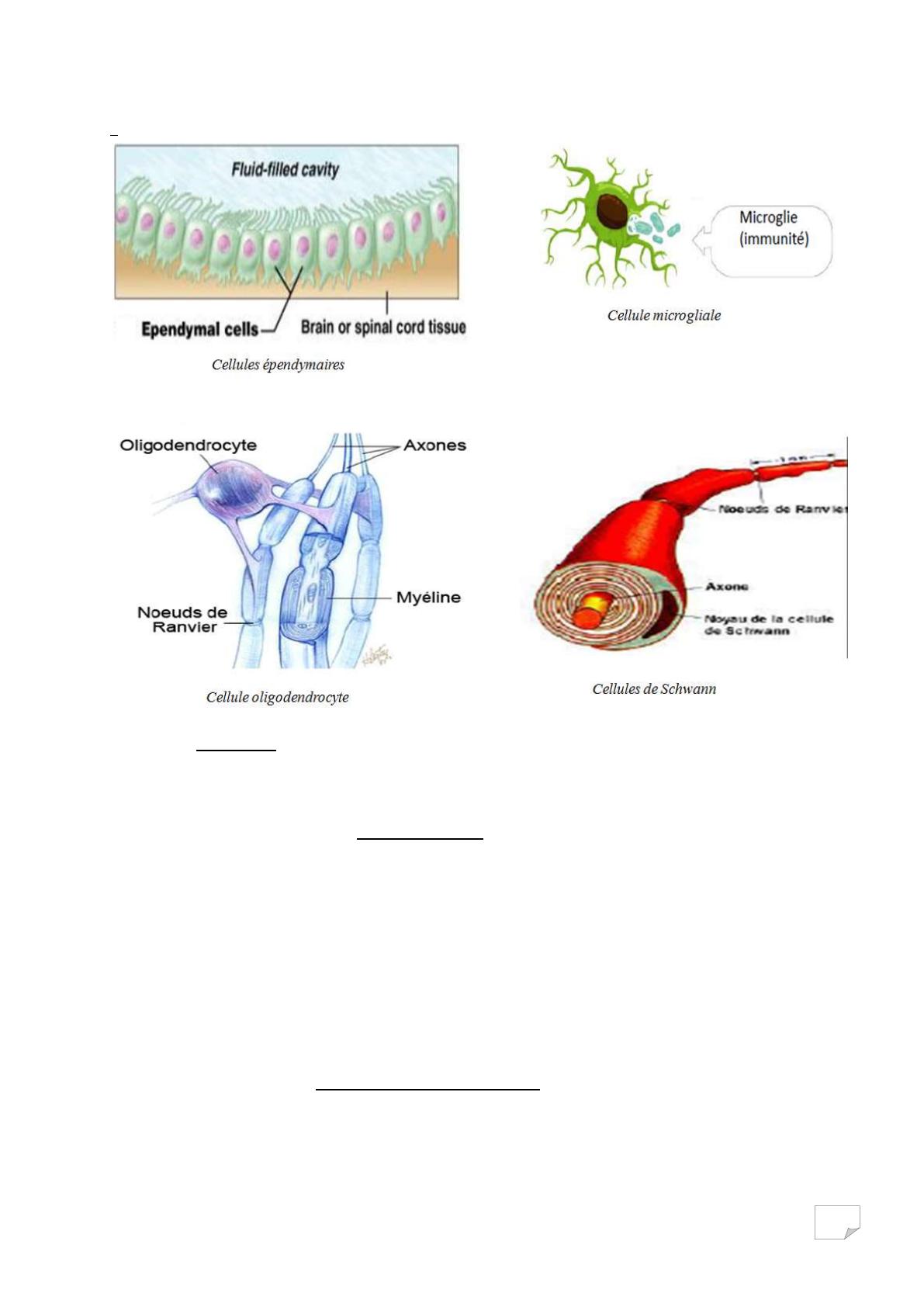
Les microgliocytes ou microglie :
De tailles réduites, avec une forte capacité de prolifération. Elles sont
capables de phagocyter les débris cellulaires et sont impliquées dans les
inflammations et les lésions.
Les épendimocytes ou les cellules épendimaires :
Cellule épithéliales tapissant l’intérieur des cavités cérébrales et de la
moelle épinière, participant à la fabrication du liquide céphalo-rachidien (LCR).
Les oligodendrocytes ou oligodendroglie :
Elles fabriquent la myéline des axones du SNC en s’enroulant autour de
ces axones et constituant (les enroulements) des isoles. Un oligodendrocyte
s’enroule autour de plusieurs axones de façon discontinue, il existe entre deux
enroulements ou gaine, un espace appelé nœud de Ranvier.
Les cellules de Schwann :
Elles fabriquent la myéline des axones du SNP et augmente la conduction
de l’influx nerveux plusieurs cellules s’enroulant autour d’un axone. Elles ont un
rôle équivalent aux oligodendrocytes et sont présentes dans le SNP.
4
!
Figure 1 : Les différentes cellules gliales (névroglie)
Les neurones :
Le neurone est l’unité structurelle et fonctionnelle du système nerveux.
Leur nombre est connu à la naissance (un certain nombre n’atteint pas la
maturité), 100 milliards environ, ils ne se reproduisent pas. Ce sont des cellules
excitables multifonctionnelles : elles reçoivent, conduisent, intègre et émettent
l’information.
La structure du neurone :
Malgré la diversité des formes, les neurones ont une structure de base
invariable : corps cellulaire, axone et terminaison nerveuse.

5
Le corps cellulaire ou soma est le centre vital de la cellule. Il possède un
noyau, un cytoplasme possédant des organites, le corps de Nissl (réticulum +
REG). Il émet des prolongements : dendrites et axone.
Les dendrites sont de fins et cours prolongements ramifiés et sont les
récepteurs de l’influx nerveux.
L’axone ou cylindraxe est prolongement unique en forme de câble, de
taille varié (dépendant de la forme du neurone) dont l’extrémité ramifié forme
l’arborisation terminal et les terminaisons renflées « boutons terminal ». Il est
entouré de deux types de gaines : la gaine de Schwann à rôle de soutien et de
nutrition et la gaine de myéline à présence facultative, est interrompue au
niveau des nœuds de Ranvier.
L’influx nerveux a un seul et unique sens : des dendrites ou soma à
l’axone et de l’axone à l’arborisation terminale.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
1
/
36
100%