Structure et grandes fonctions métaboliques des plantes La

Structure et grandes fonctions métaboliques des plantes
La transpiration
Evaporation, flux transpiratoire (gradient de Ψ) et résistance à la diffusion (cuticulaire, couche
limite, stomate)
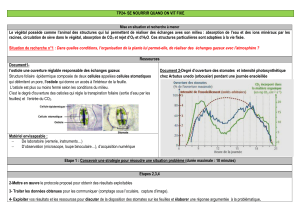
La plante perd de l’eau par évaporation au niveau des stomates des feuilles en même temps
qu’elle réalise ses échanges gazeux. Pour les plantes en C3, le rapport de transpiration (eau
perdue) / (CO2 fixé) = 500. Pour les C4 ce rapport vaut 250 et enfin pour les CAM, il est de
50. Au niveau des parois des cellules mésophyliennes, il y a un mécanisme physique
d’évaporation : l’eau passe de l’état liquide à l’état gazeux.
L’eau évaporée va diffusée, un flux transpiratoire se met en place grâce au gradient de
potentiel hydrique Ψ. Plus l’humidité augmente plus Ψ augmente et devient quasiment celui
de l’eau pure (nul). Le Ψ foliaire se situe entre – 0,8 et – 1,5 MPa. Il s’agit du gradient de Ψ
entre les feuilles et l’air. Il dépend de la résistance à la diffusion de l’eau.
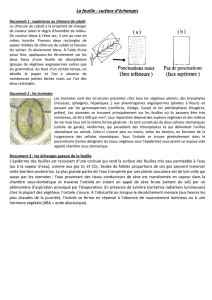
Il y a différents types de résistances : résistance cuticulaire (élevée), résistance de la couche
limite (élevée), résistance stomatique (faible). La couche limite correspond à la couche d’air
non agitée au contact de la feuille plus ou moins saturée en eau. L’air extérieur est une
atmosphère dite agitée : la résistance est proportionnelle à l’épaisseur de la couche limite.
Mouvement stomatique et contrôle de la transpiration
Influence des facteurs externes
Les stomates s’ouvrent le jour et se ferment la nuit. Chez les plantes CAM, ils sont ouverts la
nuit et fermés le jour. Quand la journée est ensoleillée, ils présentent une ouverture maximale.
A partir de midi la fermeture est d’abord lente puis elle s’accélère en fin de journée. Lors de
journées pluvieuses, ils s’ouvrent moins. De même lors d’un stress hydrique, ils ne s’ouvrent
que le matin et peu. La lumière, la baisse de CO2, le stress hydrique, le vent, la température
sont des facteurs contrôlant l’ouverture des stomates.
Mécanismes d’ouvertures et de fermetures
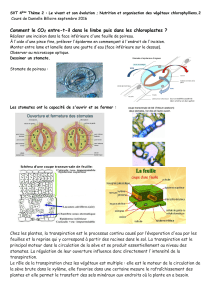
L’ouverture des stomates est provoquée par la turgescence des cellules de gardes qui
entourent l’ostiole. La paroi autour de l’ostiole est plus épaisse donc moins déformable, donc
la cellule se déforme par les parois opposées à l’ostiole qui va alors s’ouvrir.
Les phototropines sont des pigments photorécepteurs sensibles à la lumière bleue qui agit
comme stimulus dans le phénomène de turgescence. Les phototropines sont des
flavoprotéines à deux domaines LOV (sensibles à la Lumière, l’Oxygène, le Voltage). Le
vanadate, inhibiteur de la pompe à proton, fait disparaître la turgescence. A la lumière, il y a
alors activation des pompes ATPase H+ qui provoque une hyperpolarisation de la membrane,
donc une entrée d’ions. Si on ajoute du baryum, bloquant les canaux potassiques entrants, il
n’y a plus de courant. Les canaux Cl sont également impliqués. Le pH intracellulaire varie, il
y a activation de la PEPcarboxylase qui active la synthèse de malate. Une augmentation de la
concentration intracellulaire en KCl et en malate abaissent le potentiel hydrique et favorise
l’entrée d’eau dans les cellules de garde et le phénomène de turgescence.

L’ouverture des stomates est maintenue au cours de la journée par la photosynthèse. La
lumière intervient dans la photosynthèse. Les produits issus de la photosynthèse sont
transformés en saccharose dans le hyaloplasme et stocké dans la vacuole. Le potentiel
hydrique diminue donc l’eau entre et la turgescence est maintenue.
La fermeture résulte de la perte de turgescence soit par évaporation (mécanisme hydropassif)
soit par l’activité de la cellule (mécanisme hydroactif). L’acide abscissique (ABA) est
responsable de la fermeture des stomates. ABA active les canaux Ca donc du calcium entre
dans la cellule et la dépolarise. Il y a alors ouverture des canaux anioniques et notamment des
canaux K sortants. L’augmentation de la concentration intracellulaire en calcium inhibe les
ATPase H et les canaux K entrants. Il y a une sortie d’ion donc le potentiel hydrique
augmente et provoque une sortie d’eau. C’est alors la plasmolyse et la fermeture des stomates.
Lors de stress hydrique, la racine synthétise du ABA qui sera transporté par la sève brute
jusqu’aux feuilles.
1
/
2
100%