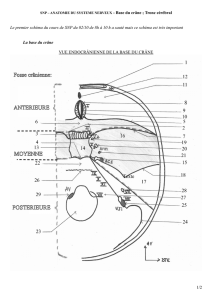
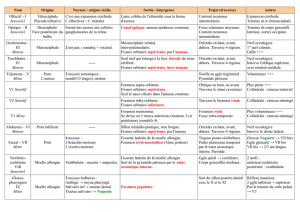
Microévolution d'une population historique : Missiminia (Soudan)
Telechargé par
jacques.picard03


Microévolution d'une population historique. Étude des
caractères discrets de la population de Missiminia (Soudan, IIIe-
VIe siècle)
In: Bulletins et Mémoires de la Société d'anthropologie de Paris, Nouvelle Série, tome 11 fascicule 1-2, 1999. pp. 1-
213.
Abstract
Skeletons of historic populations generally originate from burial grounds. They do not systematically provide a representative
sample of the population from which they derive. The most frequent factors of bias are demographic (over-or under-
representation of an age group or sex) or social (organization of graves or sectors of the cemetery according to biological and/or
family criteria). So, before starting research on the history of the population, the samples studied must be carefully defined. This
was our aim in this work on the burial ground of Missiminia (Sudan) where hundred of graves and more than 350 intact skulls
have been discovered distributed in four groups from Meroitic the Christian period... (more to read)
Résumé
Les squelettes des populations historiques proviennent généralement de cimetières. Ils ne représentent pas systématiquement
un échantillonnage correct de la population dont ils sont issus. Les facteurs de biais les plus fréquents sont démographiques (sur
ou sous-représentation d'une classe d'âge ou d'un sexe) ou sociaux (organisation sur des critères biologiques et/ou familiaux de
tombes ou de secteurs de cimetières). Dès lors, avant d'entamer une recherche sur l'histoire d'un peuplement, il faut préciser la
définition des échantillons retenus. C'est ce qui a été tenté dans cette étude portant sur la nécropole de Missiminia (Soudan) qui
a livré des centaines de tombes et plus de 350 crânes intacts provenant de 4 groupes datés de la période méroitique à la période
chrétienne... (texte à suivre)
Citer ce document / Cite this document :
Crubezy Éric, Telmon Norbert, Sevin André, Picard Jacques, Rougé Daniel, Larrouy Georges, Braga José, Ludes Bertrand,
Murail Pascal. Microévolution d'une population historique. Étude des caractères discrets de la population de Missiminia
(Soudan, IIIe-VIe siècle). In: Bulletins et Mémoires de la Société d'anthropologie de Paris, Nouvelle Série, tome 11 fascicule 1-
2, 1999. pp. 1-213.
doi : 10.3406/bmsap.1999.2537
http://www.persee.fr/web/revues/home/prescript/article/bmsap_0037-8984_1999_num_11_1_2537

Bulletins
et
Mémoires
de
la
Société
d'Anthropologie
de
Paris
1999,
nouvelle
série
-
Tome
11,
Fascicule
1-2,
pages
1
à
213.
Microévolution
d'une
Population
Historique
Étude
des
caractères
discrets
de
LA
POPULATION
DE
MlSSEVflNIA
(Soudan,
IIP-VP
siècle)
MlCRO-EVOLUTION
OF
Historic
Populations
Study
of
discrete
traits
of
the
population
of
mlssiminia
(Sudan,
III^-VI™
century
A.D.)
En
hommage
à
Madame
Ginette
Billy
et
à
Monsieur
André
Vila
Eric
Crubézy1,
Norbert
Telmon1,
André
Sevin1,
Jacques
Picard2,
Daniel
Rouge1,
Georges
Larrouy1,
José
Braga3,
Bertrand
Ludes4,
Pascal
Murail3
1.
Université
Paul
Sabatier,
UMR
8555
du
CNRS.
39
Allées
Jules
Guesde,
31
000
Toulouse,
e-mail
2.
Centre
de
Recherche
du
Service
de
Santé
des
armées
3.
Université
de
Bordeaux
I,
UMR
5809
du
CNRS
4.
Institut
de
Médecine
Légale,
Université
Louis
Pasteur,
Strasbourg,
UMR
8555
du
CNRS

Bulletins
et
Mémoires
de
la
Société
d'Anthropologie
de
Paris
1999,
nouvelle
série
-
Tome
11,
Fascicule
1-2,
pages
1
à
213.
SOMMAIRE
-
CONTENTS
Microévolution
d'une
population
historique
Etude
des
caractères
discrets
de
la
population
de
Missiminia
(Soudan,
IIP-VP
siècle)
Avant
Propos
7
Résumé
9
Summary
11
I.
—
INTRODUCTION,
HISTORIQUE
13
1.
—
Contexte
historique
17
2.
—
Contexte
anthropologique
18
IL
—
LA
NECROPOLE
DE
MISSIMINIA
21
1.
—
Situation
21
2.
—
Phases
d'utilisation
de
la
nécropole
25
3.
—
Les
sujets,
recrutement
et
démographie
30
Ш.
—
CARACTÈRES
DISCRETS
ET
MARQUEURS
GÉNÉTIQUES
:
CHOIX
ET
TRAITEMENT
37
1.
—
Définition
et
dénomination
des
caractères
discrets
37
2.
—
Étiologie
des
caractères
discrets
38
3.
—
Caractères
discrets
et
marqueurs
génétiques
42
4.
—
Choix,
codage,
démarche
suivie
en
analyse
univariée
44
5.
—
Recherche
de
regroupements
topographiques
en
analyse
univariée
48
6.
—
Démarche
suivie
en
analyse
multtvariée
55
7.
—
Études
inter-groupes
et
analyse
des
données
63
8.
—
Sélection
des
caractères
discrets
64

E.
CRUBÉZY
ETAL—
MICROÉVOLUTION
D'UNE
POPULATION
HISTORIQUE
DU
SOUDAN
IV.
—
ÉTUDE DES
CARACTÈRES
DISCRETS
67
1.
—
Les
caractères
hypostotics
67
1.1.
—
Suture
métopique
présente
(Sutura
frontaljs)
67
1.2.
—
Scissure
pétro-squameuse
postérieure
présente
(Fissura
squamomastoidea)
69
1.3.
—
Sutura
transversa
squamae
occipuaus
70
1.4.
—
Sutura
mendosa
72
7.5.
—
Suture
infra-orbitaire
présente
(Sutura
infraorbitaus)
73
1.6.
—
Traces
de
l'os
japonicum
(traces
de
la
sutura
zygomatica)
et
os
japonicum
(os
zygomaticum
bipartitum)
74
1.7.
—
Suture
incisive
visible
(Sutura
incisna)
76
1.8.
—
Trou
ovale
incomplet
(Foramen
ovale
incompletum)
77
1.9.
—
Foramen
épineux
incomplet
(Foramen
spinosum
incompletum)
78
1.10.
—
Foramen
de
Huschke
(Foramen
tympanicum)
80
1.11.
—
Facette
condyuenne
double
(Faciès
articularis
condylaris
bipartita)
.
81
1.12.
—
Fossette
naviculaire
81
1.13.
—
Fossette
pharyngienne
84
1.14.
—
Agénesiedela
troisième
molaire
supérieure
85
1.15.
—
Caractères
hypostotics
:
récapitulation
85
2.
—
Les
caractères
hyperostotics
88
2.1.
—
Torus
palatin
(Torus
palatinus)
88
2.2.
—
Épine
throchléaire
(Spina
throchlearis)
91
2.3.
—
Épine
ptérygo-alaire
(Spina
pterygo-alaris)
91
2.4.
—
Pont
ptérygo-alaire
(Ponticulus
pterygoalaris)
92
2.5.
—
Tubercule
précondyuen
(Tuberculum
praecondylare)
95
2.6.
—
Canal
hypoglosse
bipartite
ou
multiple
(Canaus
hypoglossi
^
bipartite
ou
multiple)
96
2.7.
—
Canal
condyuen
intermédiaire
(Canaus
condylaris
intermedius)
96
2.8.
—
Processus
para-condyuen
(Processus
paracondylaris)
100
2.9.
—
Récapitulation
:
caractères
hyperostotics
101
3.
—
os
suturaires,
fontanell
aires,
surnumeraires
104
3.1.
—
os
suturaire
coronal
(ossa
suturae
coronaus)
106
3.2.
—
os
suturaire
sagittal
(ossa
suturae
sagittaus)
106
3.3.
—
os
suturaire
lambdoïde
(ossa
suturae
lambdoïdeae)
106
3.4.
—
Préinterpariétal
(Os
preinterparietale)
107
3.5.
—
os
au
ptérion
(os
pteril)
109
3.6.
—
Articulation
fronto-temporale
(Sutura
frontotemporaus)
110
3.7.
—
Os
À
l'incisure
pariétale
(Os
incisurae
parietaus)
112
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
1
/
208
100%