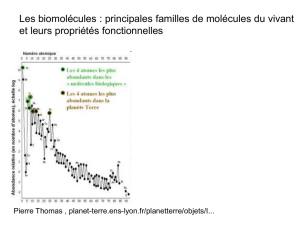
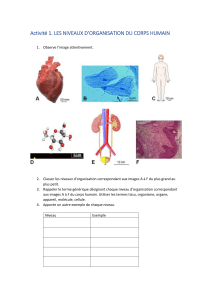
télécharger le cours - e

PARTIE 1 : ASPECTS STRUCTURAUX
I. Généralités
Les sucres représentent l’un des composants importants des
organismes vivants aussi bien que les lipides. Les glucides
interviennent dans la structure cellulaire et tissulaire et pour
certains d’entre eux, ils constituent une réserve énergétique.
Les glucides, du nom latin «glucis», aussi appelés sucres,
évoquant leur saveur sucrée. Il existe une grande diversité des
molécules dont l’assemblage sous forme de chaine complexe,
linéaire ou ramifiée, constitue les polysaccharides. Ces structures complexes constituent au niveau des
être vivants les structures biologiques, impliquées dans de nombreux phénomènes de reconnaissance
intermoléculaire mais également intercellulaire.
A. Composition des sucres
Les sucres sont également appelés hydrates de carbone (dénomination à ne pas utiliser). Les glucides
possèdent des atomes de Carbone, d’Hydrogène et d’Oxygène.
De nombreux glucides ont des formules chimiques brutes de la forme : Cn (H2O)n , qui suggère que les
glucides sont des hydrates de carbone : c’est le cas du glucose (poly). Dénomination à éviter car tous
les glucides ne se conforment pas à cette formule brute, certains contiennent en plus :!
Azote N (ester azoté), Phosphore P (ester phosphorique), Soufre S (ester sulfurique)
Cette dénomination est également fausse car certains composés qui se conforment pourtant à cette
formule brute n’appartiennent pas à la famille des glucides (comme l’acide acétique).
B. Classification des glucides
Ils sont classés en 3 catégories en fonction de leur tailles.
-Les Monosaccharides (oses simples) : Constitués d’une seule unité polyhydroxylée et peuvent
posséder une fonction de type aldéhydique (D-glucose) ou de type cétonique (D-fructose).!
Exemple : Le glucose est le monosaccharide le plus abondant dans la nature et possède 6 atome de
carbone
-Les Oligosaccharides : Ce sont de très courtes chaines composées de quelques unités
monosaccharidiques (minimum 2, maximum 10).!
Exemple : disaccharides (lactose, maltose, saccharose)
—————————————!
sur 1 10
LES GLUCIDES
UE1

-Les Polysaccharides : Ce sont de longues chaînes constituées de plusieurs centaines/milliers
d’unités monosaccharidiques. Certains sont sous forme linéaire (cellulose) ou de chaînes ramifiées
(glycogène). Il continent de 100 à + de 1000 unités monosaccharidiques.!
II. Les monosaccharides!
Ils sont appelés sucres simples, car ils sont non hydrolysables (ne peuvent pas être scindés) et portent
la plupart du temps entre 3 et 7 atomes de carbone (C). Leur squelette est constitué par une chaine
carbonée non ramifiée dans laquelle tous les atomes de C sont reliés entre eux par des liaison simples.
Ce sont des polyols (portent 2 fonctions alcool), dont l’une est une fonction alcool primaire et l’autre
une fonction réductrice carbonylé soit de type aldéhydique soit de type cétonique.
Le fait qu’ils comportent le groupement OH fait que ce sont des molécules très hydrophiles (solubles
dans l’eau).
⇒ 3C = triose 4C = tétrose 5C = pentose 6C = exose 7C = heptose ⇐
•Les trioses : 3C
Ce sont les plus petits composés répondant à la définition des oses. Il en existe que 2 :!
- D-glycéraldéhyde : fonction aldéhyde!
- D-hydroxyacétone : fonction cétone
Il faut noter que les deux composés, sous leur forme phosphorylée, correspondent à une étape clé qui
intervient en particulier dans la voie de la glycolyse. !
Fructose 1.6 diphosphate → Glycéraldéhyde P + Dihydroxyacétone P
On appelle isomères les composés ayant la même formule brute (pour le triose, C3H6O3), mais des
formules développées différentes; c’est le cas du glycéraldéhyde et du dihydroxyacétone qui se
distinguent via la fonction aldéhyde ou la fonction cétone portée : on parle donc d’isomérie de
fonction.
En plus de la longueur de la chaine carbonée, et de la nature de la fonction réductrice, les oses peuvent
se différencier par la position dans l’espace des différents groupements hydroxylés qu’ils possèdent.
→ On aborde la notion de C asymétrique, il s’agit d’un Carbone lié à 4 groupements différents, et en
fonction de la position dans l’espace de la molécule, il peut soit être à droite, soit être à gauche. Par
convention, l’hydroxyde qui se trouve à droite est nommé D-Glycéraldéhyde. À gauche, on parle de
L-Glycéraldéhyde. On parle alors de stéréoisomérie ou isomérie optique. Leur disposition dans
l’espace sera différente en fonction de la position des groupements hydroxyles. !
Pour les aldoses, ce C1 en haut correspond au
carbone le plus oxydé.!
Dans le cas des fonctions aldéhydes, il est donc situé
en haut.
—————————————!
sur 2 10

Les chaines carbonées où le groupement OH est à droite sont appelées de forme D et à gauche sont de
la forme L.
Ces deux formes sont des énantiomères qui possèdent des propriétés physiques et chimiques
identiques.!
!
In vivo, certains enzymes présentent une stéréospécificité très étroite vis à vis de leur substrat et sont
capables de distinguer soit l’un ou soit l’autre des énantiomères. Elles vont soit catalyser le D-
Glycéraldéhyde, soit le L-Glycéraldéhyde.
En revanche, en solutions, ils ont des pouvoirs différents. En effet, en solution, les formes
énantiomères des molécules portant un C asymétrique (avec 4 groupements différents) et la position
des groupements hydroxyles qui se trouvent dans le plan soit à droite, soit à gauche, va permettre
effectivement d’avoir une propriété optique différente. Chacun de ces molécules, soit le D-
Glycéraldéhyde ou le L-Glycéraldéhyde, vont dévier une lumière polarisée.
La lumière est en général polychromatique, et grâce à un filtre, elle va devenir monochromatique et
donc polarisée, et lorsqu’elle traverse cette solution de sucres, elle peut la dévier d’un angle ɑ vers la
gauche, donc dans le sens inverse des aiguilles d’une montre, on dira alors par convention que la
substance est lévogyre. Le phénomène inverse où certaines solutions vont dévier cette lumière
polarisée vers la droite, soit dans le sens des aiguille d’une montre, on dira par convention que cette
substance est dextrogyre.!
!
Un mélange équimolaire de 2 énantiomères, qui sont dextrogyres ou lévogyres est optiquement
inactive. Ce sont des mélanges racémiques.!
!
→ Concernant le Dihydroxyacétone : il ne possède pas de carbones asymétriques. Cette molécule sera
optiquement inactive.
La configuration et le pouvoir rotatoire sont indépendants. Il n’existe aucun lien entre la forme D ou L
(position droite ou gauche du groupement -OH du premier C*) et le sens de déviation droite ou gauche
du plan de polarisation de lumière. !
!
Certains oses de type D seront dextrogyres, d’autre seront lévogyres.
Exemple : le D-glucose est dextrogyre (+), mais le D-Fructose est
lévogyre (-) !
•Pour les autres oses : nC > 3
Il y a ajout d’un nouveau Carbone asymétrique.
Nombre de stéréoisomères: N = 2n n = nombre de carbones asymétriques
À chaque addition d’un C asymétrique, il existe 2 possibilités : le groupement hydroxyle du nouveau
carbone peut être à droite ou à gauche de l’axe de la chaine carbonée. On obtient un nombre total de
stéréoisomère qui sera égal au nombre de combinaison possible de chaque atome de C*.
—————————————!
sur 3 10
D-Glucose!
Dextrogyre (+)
D-Fructose!
Levogyre (-)

Exemple de l’aldohexose : N= 24 = 16 stéréoisimères (4 C*)
On représente uniquement les carbones au niveau des intersections dans les représentation de Fisher et
les traits horizontaux représentent la localisation des OH. Les hydrogènes en face ne sont pas
représentés.
Nomenclature des oses : elle est relative. Tous les sucres qui sont préfixés par la lettre D ou L, font
référence au triose de base qui est le D- ou L-glycéraldéhyde. !
Ce préfixe sera suivi de la nature de pouvoir rotatoire de la molécule qui est soit « + » (droite) soit !
« - » (gauche).
D-éythrose et D-thréose sont énantionmères.
Tous les oses qui comporteront et qui font référence à cette molécule
appartiennent à la séries des D-oses.
Un se distingue : le D-ribose (C5). C’est un composant de l’ARN utilisé dans la transcription
génétique et est apparenté au D-désoxyribose (composant de l’ADN).
Filiation aldoses - C6 - série D
6 Carbones, dont 4 Carbones asymétriques. Il y
a 16 stéréosiomères, 8 de la série D et 8 de la !
série L.
Tous ceux de la série D font références au !
D-glycéraldéhyde.
→ Quand 2 monosaccharides ne diffèrent que par la configuration d’un seul atome de carbone
asymétrique, on dit qu’ils sont épimères l’un par rapport à l’autre : ils constituent un couple
d’épimères
→ Le D-glucose et le D-mannose sont épimères en C2.!
→ Le D-glucose et le D-galactose sont épimères en C4.
➡Les cétoses (comportent une fonction cétone):
Représentants: les dihydroxyacétones
La nomenclature est définie par rapport à la position de l’hydroxyle porté
par le C* voisin de la fonction primaire la plus éloignée de la fonction
cétone en référence au dihydroxyacétone.
Absence de carbone asymétrique pour le dihydroxyacétone ! !
!
Les cétoses présent un C* de moins par rapport aux aldoses.!
—————————————!
sur 4 10

Filiation cétoses - C3 à C6 - série D
Le plus connu est le D-fructose qui ne comporte que 3C* et qui est le constituant principal du
saccharose.!
Le fait que les cétoses possèdent 1 C* en moins entraine un nombre de stéréoisomères qui sera 2 fois
moindre que celui des aldoses.
→ Il existe 4 aldotétroses mais seulement 2 cétotétroses!
→ Il existe 16 aldohexoses mais seulement 8 cétohexoses
Oses simples d'intérêt biologique
Hormis de rares exception, les oses naturels et leurs dérivés sont de la série D.
-Trioses: C3 → D-Glycéraldéhyde et ses dérivés phosphorylés que l’on peut retrouver dans les
étapes de la glycolyse. Très présents dans la nature.
-Pentoses: C5 → D-Ribose et son dérivé le D-2 désoxyribose sont des composants des acides
nucléiques.
-Hexoses: C6 donc 4C*. On retrouve le :
→ D(+) Glucose : molécule carburant et réserve énergétique, hydrosoluble dans les liquides
biologiques. Dans les végétaux, le D-glycopyranose se retrouve dans l’amidon et chez les
animaux dans le glycogène. Présent aussi dans le miel.
→ D(+) Galactoses : ose le plus répandu après le glucose, constituant du lactose (lait des
mammifères) et on le retrouve combiné dans certains oligosydes hétérocycles ou encore des GP.
→ D(-) Fructose : rare sucre cétonique naturel, on le retrouve à l’état naturel dans les fruits, le
miel auquel il donne sa consistance à cause de sa cristallisation relativement difficile et surtout,
il rentre dans la composition du saccharose.
→ D(+) Mannose : présent dans le mannane et dans les écorces d’orange!
Phénomène de mutarotation et anomère (propriétés physiques des oses)
Lorsque l’on met du D(+) Glucose en solution, on va avoir un
pouvoir rotatoire qui va varier au cours du temps. On retrouve
deux types de formes en solution, les formes ɑ et les formes β.
Ces deux types de formes, qui sont liées au sucre en solution,
sont dites anomères. !
!
Cela suggère en fait que le D(+) Glucose en solution a un
centre chiral, donc un carbone asymétrique supplémentaire et
que lorsque l’équilibre est atteint, les deux formes ɑ et β sont
présentes en solution dans un rapport respectif de 1/3 et 2/3.
!
—————————————!
sur 5 10
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%