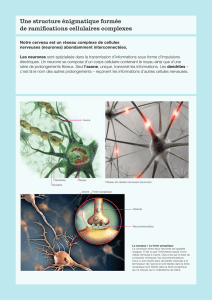
La formation et l`élimination des synapses

La formation et
l’élimination des synapses
Développement et plasticité
neuronale
Images tirées de « Fundamental Neuroscience », ©Academic Press, 1999
Traduction et présentation, © Claude Messier, 1999

Formation des synapses...
• Le premier contact de l ’axone avec sa cellule-
cible initie une série d ’interactions impliquant
un échange complexe de signaux entre
l ’axone et la cellule-cible qui mènera à la
formation d ’une synapse.
• Le système nerveux en développement
fabrique des synapses en excès (en gros deux
fois plus qu ’il n ’en restera à l ’âge adulte.

Et élimination des synapses...
• L ’élimination des synapses est un élément
crucial pour le raffinement fonctionnel des
circuits neuronaux.
• Les axones présynaptiques entrent en
compétition pour une connexion avec une
cellule-cible post-synaptique.
• L ’axone perdant retire ses synapses et
quelquefois (pas toujours) la cellule dont est
issu l ’axone est « tuée ».

Résumé
• C ’est donc la combinaison de ces deux
processus développementaux que les patrons
spécifiques et finals des connections adultes
émergent durant le développement.

La plupart des informations sur les mécanismes
de la formation des synapses nous viennent de
l ’étude de la jonction neuromusculaire
Axone d’un motoneurone cellule de schwann
Cellule musculaire
replis membranaires
post-synaptiques
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
1
/
35
100%