Organogenèse des membres

1
Organogenèse des membres
I. Formation du bourgeon de membre de vertébré
A-Organisation des membres chez les vertébrés
B- Morphogenèse du membre de vertébré
C- Spécification du territoire du membre
D- Induction du bourgeon de membre
II. Mise en place des axes de polarité du membre
A- Régionalisation proximo-distale
A.1. Le bourrelet apical ectodermique (AER: «!Apical ectodermal ridge!»)
A.2. La zone de progression («!progress zone!»)
A.3. Expression régionalisée des gènes Hox
B- Régionalisation antéro-postérieure
B.1. La zone d’activité polarisante (ZPA)
B.2. Shh est la molécule de!signalisation produite par la ZPA
C- Régionalisation dorso-ventrale
III. Spécification de l’identité des membres, antérieur versus postérieur
A- Spécification par les gènes Hox
B- Spécification par les facteurs de transcription Tbx
IV. Différenciation tissulaire dans le membre
A- Le squelette
B- Les muscles
C- L’innervation
D- la vascularisation
E- L’individualisation des doigts nécessite une apoptose morphogène
CAPES 2006-2007 cours: Christine Laclef

2
A- Organisation des membres de vertébrés
I – Formation du bourgeon de membre de vertébré
Différents axes de polarité bien visible au niveau de la main

3
Différents axes de polarité visibles
dans l’aile de poulet
A- Organisation des membres de vertébrés

4
Diversité de forme des éléments squelettiques,
mais conservation des subdivisions:
- Stylopode: élément proximal
- Zeugopode: élément médian
- Autopode: élément distal
A- Organisation des membres de vertébrés

5
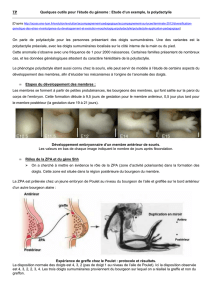
Morphogenèse du membre
embryons humains
29 jours 32 jours
M: mésenchyme
E: ectoderme
AER
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
1
/
48
100%