12. Les plantes et l`eau - Agence de l`Eau Seine Normandie

69
12. Les plantes et l’eau
Les plantes sont constituées de 60 à 90 % d’eau. Leur vie en dépend, toujours pour leur
physiologie, souvent pour leur biologie. L’évolution leur a permis de sortir de l’eau, puis de
s’adapter à tous les milieux terrestres, des plus secs aux plus humides.
1. L’eau physiologique
L’eau entre et circule dans la plante, elle peut aussi en sortir. Ces étapes sont
interdépendantes, chacune a un rôle important. Dans les cellules où sont présents les
chloroplastes, la première réaction de la photosynthèse est la photolyse de l’eau. Dans
toutes les cellules vivantes, les mitochondries permettent la respiration dont la dernière
réaction produit de l’eau.
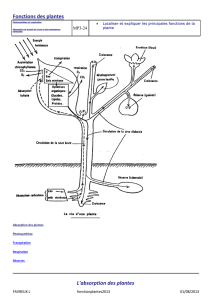
a. Le transfert de l’eau dans la plante
1. Ce qui entre
Chez les plantes à racines-tiges-feuilles (les Cormophytes), l’eau entre par les racines
suivant la loi de l’osmose en fonction de la surface de contact des poils absorbants ou
des mycorhizes avec l’eau du sol.
L’eau passe du milieu où elle est relativement plus abondante, c’est-à-dire le milieu
le moins concentré en substances dissoutes, vers le milieu où elle est relativement
moins abondante, c’est-à-dire le milieu le plus concentré en substances dissoutes.
La grande diversité des qualités physiques et chimiques des sols infl uence très
fortement ce passage. Prenons deux cas extrêmes : le sable étant perméable
ne permettra pas aux plantes d’absorber l’eau très longtemps ; les sols argileux
retiennent bien l’eau, ce qui est favorable aux plantes, mais ils peuvent se saturer
et devenir imperméables et anoxiques ou s’assécher complètement et se craqueler.
2. Ce qui circule
Pour que l’eau circule dans la plante, il faut que les stomates (photo 1) soient ouverts
au niveau des feuilles, qu’il y ait donc transpiration. C’est un système de pompe,
l’eau est aspirée des racines vers les feuilles.
Cette eau et les sels minéraux dissous circulent dans les vaisseaux du bois ou
xylème : c’est la sève brute (photo 2). Après la photosynthèse, la sève élaborée
amenant les produits carbonés aux tissus circule en sens inverse dans les vaisseaux
du liber ou phloème.
L’eau est présente également dans les méats cellulaires et dans les lacunes aquifères
(photo 3).
1. Face inférieure d’une feuille
de laurier rose
2. Coupe transversale
d’une racine d’iris 3. Coupe transversale
de pétiole de nénuphar
Stomate au fond d’une crypte pilifère Vaisseau de bois
Vaisseau de liber Aérenchyme
Lacune aérifère
Lacune aquifère

70
3. Ce qui sort
Il existe des barrières effi caces qui protègent la
plante contre la dessiccation, la plus superfi cielle
étant la cuticule très hydrophobe (photo 4).
La transpiration se fait par les stomates. Il existe
une régulation dépendant de la concentration en
C02, en relation avec le type métabolique, C3 ou C4
ou CAM (cf. ci-après). L’humidité de l’air infl uence
également l’ouverture des stomates. La « couche
limite », c’est-à-dire l’accumulation de vapeur
d’eau au contact même de la feuille, offre une
résistance à la sortie de l’eau. Le vent tend à
supprimer cette couche-limite, donc à favoriser
la transpiration (sauf adaptation au xérophytisme
comme la pilosité, voir photo 5).
b. L’eau métabolique
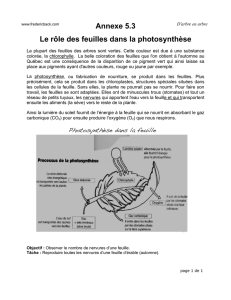
1. La photosynthèse
Les végétaux verts sont les producteurs primaires utilisant le dioxyde de carbone
atmosphérique pour réaliser les substances carbonées utilisées dans la chaîne
trophique. Le CO2 pénètre par les stomates des feuilles et arrive jusqu’aux
chloroplastes des cellules.
La réaction globale est connue sous la forme :
12H2*O + 6CO2 C6H12O6 + 6*O2 + 6H2O
Le marquage radioactif de l’oxygène de l’eau permet de constater que celle-ci est
dissociée et est à l’origine de l’oxygène libéré.
Cette dissociation est la photolyse de l’eau. La lumière agit sur la chlorophylle,
l’oxygène est libéré, l’hydrogène est pris en charge par des transporteurs chimiques
et sera intégré avec le dioxyde de carbone dans une chaîne métabolique qui aboutira
à la formation du glucose puis de l’amidon. Cette intégration du CO2 se fait chez
la majorité des plantes sur un sucre à cinq carbones et aboutit à la formation de
deux acides à trois carbones, c’est une photosynthèse en C3. Toutes les réactions
chimiques sont catalysées par des enzymes.
2. La respiration
Alors que la photosynthèse est liée à la présence de chlorophylle dans les
chloroplastes des végétaux verts, la respiration est nécessaire à la vie de toute
cellule eucaryote.
La réaction globale se présente ainsi :
C6H12O6 + 6*O2 + 6H2O 6CO2 + 12H2*O
Le marquage radioactif de l’oxygène montre que celui-ci se retrouve intégralement
dans l’eau formée.
Cette réaction se fait en trois étapes dans chaque cellule. La première étape se
4. Coupe transversale de laurier rose
5. Medicago marina

71
fait dans le hyaloplasme : c’est la glycolyse ; le glucose perd des H qui se fi xent sur
des transporteurs chimiques, il se forme un produit intermédiaire. Les deuxième
et troisième étapes se font dans les mitochondries : ce produit perd du CO2 qui
est libéré et des H qui sont fi xés sur des transporteurs. La respiration se termine
par l’intervention de l’O2 qui provoque la libération de l’H2 des transporteurs avec,
simultanément, et formation de H2O et production importante d’ATP, molécule
énergétique utilisée par la cellule dans différentes fonctions.
c. L’eau et la biologie
1. Dissémination par l’eau des produits liés à la reproduction
Chez les Thallophytes, les zygotes et les spores sont disséminés par l’eau,
qu’ils soient mobiles (planozygotes, planospores) ou immobiles (aplanozygotes,
aplanospores). Ceci est vrai pour toutes les spores, celles issues de la méiose à
l’origine des gamétophytes et celles issues de mitoses à l’origine de multiplication
végétative.
Chez les Embryophytes, tels que Mousses et Fougères, les gamètes mâles fl agellés
(planogamètes) rejoignent les gamètes femelles (oosphères) par voie aquatique.
Chez les Angiospermes revenus à la vie aquatique, plantes hydrophiles, les fruits
sont transportés et dispersés par l’eau, c’est l’hydrochorie.
2. Intervention mécanique de l’eau
Quand l’air ambiant devient suffi samment sec,
l’évaporation de l’eau des cellules des anneaux
mécaniques des sporanges des Fougères (photo
6) et des sacs polliniques des Phanérogames
crée une traction qui provoque l’ouverture de ces
organes, ce qui libère les spores des unes et le
pollen des autres.
L’équilibre hydrique de la plante est fonction
de nombreux facteurs afférant aux différents
types de sol, aux caractéristiques des nombreux
climats et de leur variation au cours de l’année, aux stratégies d’adaptation de la
plante quand elles sont mises en place par l’évolution. L’eau est un facteur limitant
pour toutes les plantes à des degrés différents, le stress hydrique est plus ou moins
rapidement atteint.
2. L’environnement hydrique des plantes
Au cours de l’évolution, l’eau a été le seul milieu de vie des êtres vivants pendant
3,4 milliards d’années. Puis les premiers Embryophytes ont trouvé les solutions
nécessaires pour en sortir il y a 400 millions d’années. Ces végétaux ont ensuite
colonisé les écosystèmes les plus variés.
a. Les contraintes de la vie terrestre
Il a fallu pour la plante en milieu aérien gérer l’eau, la récupérer, la faire circuler,
limiter ou tolérer les pertes. Les racines sont liées à l’existence d’un appareil
vasculaire constitué de vaisseaux de bois véhiculant la sève brute et de vaisseaux de
liber véhiculant la sève élaborée.
6. Sporange de fougère
Anneau mécanique d’un sporange
Sporange ouvert
Quelques spores non libérées

72
Le maintien des végétaux hors de l’eau nécessite des tissus de soutien dont le bois et le
sclérenchyme, la lignine étant la matière constitutive la plus effi cace dans le port des
Cormophytes.
L’autre problème à résoudre est celui de la fécondation. Chez les Thallophytes, dans
l’eau, l’hydrogamie est de règle. Qu’ils soient mobiles ou non, les gamètes mâles et
parfois les gamètes femelles sont libérés et la fécondation se fait donc dans l’eau qui
reste le milieu de vie de l’œuf qui s’y développera sans autre protection. À partir de
leur sortie de l’eau, les Embryophytes évoluent en passant de l’hydrogamie, gamètes
mâles mobiles se déplaçant (chimiotactisme positif) vers les gamètes femelles fi xes,
à la siphonogamie, gamètes mâles non mobiles apportés jusqu’aux gamètes femelles
par le tube pollinique.
Dès leur sortie de l’eau, les végétaux
devaient protéger leurs embryons
contre la dessiccation. Ils ont inventé
pour cela l’archégone (photo 7), c’est-à-
dire le gamétange femelle, protecteur
de l’oosphère (cellule sexuelle femelle)
puis de l’œuf après la fécondation et du
jeune embryon. Ces végétaux sont les
Archégoniates.
Embryophytes, Cormophytes, Archégoniates
recouvrent les mêmes végétaux, ceux qui
sont sortis de l’eau, même s’ils s’y sont
réadaptés.
Cette évolution aboutit à l’ovule qui devient
la graine à téguments plus ou moins
imperméables, puis à l’ovaire qui devient le
fruit adapté à une dispersion hydrophile, anémophile ou zoophile. Les fruits charnus
gorgés d’eau et de sucre sont particulièrement attractifs pour les animaux qui les
repèrent très vite grâce à leur couleur. Ce groupe végétal terme actuellement abouti
de l’évolution est celui des Angiospermes.
b. Le xérophytisme
Un végétal qui vit en milieu sec est un xérophyte. L’aridité du milieu peut être due soit
au climat, soit au sol. L’exemple du Sahara vient immédiatement à l’esprit, de même
que l’Atacama au Chili.
Dans nos régions plus tempérées, c’est surtout le sol qui est en cause : dunes (photo 8),
cordons littoraux (photo 9), rochers granitiques (photo 10), falaises (photo 11), vases
salées (photo 12) par exemple. Le sel apporté par les embruns est un facteur de
dessèchement supplémentaire.
7. Archégones de mousse
8. Dune de l’Amélie (33) 9. Le sillon de Talbert (22) 10. Crithmum maritimum sur les
rochers de Plougrescant (22)

73
Les caractères adaptatifs qui se sont inscrits
dans l’évolution sont parfois observables sur le
végétal. Prenons l’exemple du cactus (photo 13) :
la tige peut faire des réserves d’eau, les feuilles
transformées en épines évitent à la plante une
trop grande perte d’eau par transpiration.
Il existe d’autres caractères morphologiques mais
aussi anatomiques et physiologiques.
Voici les différentes stratégies de résistance à la
sécheresse :
• Les plantes ont acquis la faculté de se développer à
grande vitesse, c’est le cas des éphémérophytes des
déserts. Trois semaines suffi sent de la germination
à la fructifi cation.
• Il existe deux formes opposées de racines en
fonction de la présence de l’eau disponible. Les unes
sont pivotantes et peuvent s’enfoncer jusqu’aux
nappes phréatiques, à 40 m pour les acacias du
Sahara. Les autres sont traçantes dans toutes les
directions et occupent un espace superfi ciel
important permettant une très rapide absorption
de l’eau avant qu’elle ne s’infi ltre rapidement dans le
sable, c’est le cas de l’oyat sur nos dunes littorales.
• De nombreuses familles de plantes dites « grasses »
constituent des réserves d’eau (et non de lipides !).
Les plus connues sont les Cactacées des déserts
américains ; chez nous ce sont les Crassulacées
(photo 14) qui sont les plus représentatives. Notons
aussi Crithmum chez les Apiacées (photo 10),
Salicornia chez les Chénopodiacées (photo 12).
• Les adaptations consistant à réduire les pertes d’eau
sont :
- morphologiques : elles concernent surtout les
feuilles dont la surface est minime voire nulle,
réduisant ainsi la transpiration.
Si elles sont normalement développées, elles
peuvent s’enrouler par temps sec.
On remarque que chez les Cactacées la
forme sub-hémisphérique à ovoïde est assez
commune, c’est en effet la sphère qui possède le
plus faible rapport Surface sur Volume, limitant
ainsi de façon optimale la transpiration.
11. Falaise du Cap de La Hève (76)
12. Estuaire du Jaudy (22)
13. Cactus
14. Sedum refl exum sur falaise
calcaire
 6
7
8
6
7
8
1
/
8
100%