Muscle lisse bronchique et inflammation : article de pneumologie

MISE AU POINT
6
La Lettre du Pneumologue - n° 2 - décembre 1998
e muscle lisse des voies aériennes est l’un des princi-
paux effecteurs de la réactivité bronchique, c’est-à-
dire de la capacité des voies aériennes à réduire leur
diamètre en réponse à de nombreux stimuli de nature phy-
sique, chimique ou pharmacologique. Il a longtemps été consi-
déré comme un acteur passif dans l’inflammation bronchique,
se contractant en réponse à des substances pro-inflammatoires
et se relaxant en réponse à l’adrénaline circulante ou à des neu-
rotransmetteurs relaxants. Plus récemment, de nombreuses
publications ont montré que le muscle lisse pouvait, en plus de
son activité contractile, être l’objet de transformations structu-
rales, hyperplasie ou hypertrophie, qui contribuent au rétrécis-
sement des voies aériennes et à l’hyperréactivité bronchique.
Enfin, il apparaît qu’en outre, le muscle lisse pourrait avoir un
rôle actif dans la synthèse et la libération de médiateurs, aussi
bien pro-inflammatoires que bronchoconstricteurs.
Après un rappel de la structure, les fonctions contractiles et
sécrétrices du muscle lisse seront envisagées.
STRUCTURE ET ULTRASTRUCTURE
Le muscle des voies aériennes est réparti tout le long de l’arbre
trachéobronchique, depuis la trachée jusqu’aux bronchioles
terminales (1). Au niveau de la trachée, il est organisé en fais-
ceaux transversaux à la partie postérieure. Au fur et à mesure
de la réduction du diamètre bronchique, le tissu musculaire
prédomine et les faisceaux sont organisés en une hélice dont le
pas augmente dans les bronches les plus distales. Les cellules
musculaires lisses des voies aériennes sont allongées, avec un
noyau central. Elles contiennent un appareil de Golgi près du
noyau et des lysosomes. Les mitochondries sont aussi locali-
sées près du noyau. Elles sont associées à des granules de gly-
cogène et à des vésicules de pinocytose. Ces mitochondries
sont présentes en faible quantité et permettent une production
d’ATP équivalant à environ 10 % de celle du muscle squelet-
tique. Le réticulum sarcoplasmique paraît peu développé et
représente seulement 2 à 5 % du volume cellulaire total, bien
qu’il ait un rôle fonctionnel important.
CONTRÔLE
(figure 1)
Deux types de messagers extracellulaires contrôlent la contrac-
tion du muscle lisse des voies aériennes grâce à une interaction
avec des récepteurs spécifiques présents sur la membrane cel-
lulaire musculaire : les neurotransmetteurs du système nerveux
autonome et les médiateurs participant au phénomène inflam-
matoire.
Neurotransmetteurs
L’innervation cholinergique constitue la commande broncho-
constrictive principale. L’acétylcholine contracte le muscle
bronchique par interaction avec des récepteurs muscariniques
dont cinq sous-types (M1-M5) ont été clonés. C’est le sous-
Le muscle lisse bronchique
Une nouvelle cellule pro-inflammatoire ?
●
M. Molimard*, M. Tunon de Lara**, R. Marthan***
* Laboratoire de pharmacologie respiratoire, CHU de Bordeaux ; INSERM (CRI
9806) : physiopathologie de la réactivité bronchique et vasculaire.
** Service des maladies respiratoires, CHU de Bordeaux ; INSERM (CRI 9806) :
physiopathologie de la réactivité bronchique et vasculaire.
*** Laboratoire de physiologie cellulaire respiratoire, université Victor-
Segalen, Bordeaux-II ; INSERM (CRI 9806) : physiopathologie de la réactivité
bronchique et vasculaire.
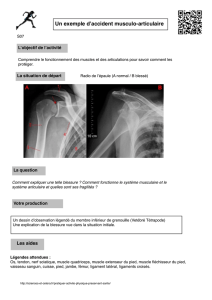
L
Neurotransmetteurs
MŽdiateurs
CGRP
ACh AD
M2
B2H1
β2
M3
NK1
NK2
VIP
LT PG Bk Hist ET NO
?
SP
NKA
NAD
ƒicosano•des
Cellule musculaire lisse des voies aŽriennes
Contraction
Relaxation
Figure 1. Principaux messagers extracellulaires impliqués dans la
contraction ou la relaxation du muscle lisse bronchique.
ACh : acétylcholine ; AD : adrénaline circulante ; Bk : bradykinine ;
B2 : récepteur B2 à la bradykinine ; CGRP : calcitonin gene-related
peptide ; ET: endothéline ; ß2 : récepteur ß2-adrénergique ; H1 :
récepteur H1 à l’histamine ; Hist : histamine ; LT : leucotriènes ; M :
récepteur muscarinique ; NAD : noradrénaline ; NKA : neurokinine A ;
NK1, NK2 : récepteurs aux neurokinines de type 1 et de type 2 ; NO :
monoxyde d’azote ; PG : prostaglandines ; SP : substance P ; VIP :
vaso-intestinal peptide.

type M3, présent sur la membrane des cellules musculaires
lisses bronchiques humaines, qui est à l’origine de la contrac-
tion du muscle lisse induite par l’acétylcholine (2, 3). Cepen-
dant, le sous-type M2 représente 80 % des récepteurs muscari-
niques présents à la surface de la cellule musculaire lisse et
pourrait avoir un rôle inhibiteur de la relaxation induite par les
agonistes ß2-adrénergiques (4, 5).
L’innervation adrénergique directe du muscle bronchique est
pauvre chez l’homme, ce qui contraste avec la densité des
ß-récepteurs présents dans le parenchyme (6). La membrane
cellulaire du muscle bronchique humain présente exclusive-
ment le récepteur β2, dont l’affinité pour la neurohormone
adrénaline est plus grande que pour le neurotransmetteur nora-
drénaline. L’adrénaline circulante est donc le principal agoniste
physiologique du récepteur ß2-adrénergique bronchique (7).
L’innervation non adrénergique non cholinergique (NANC)
contrôle la contractilité du muscle lisse trachéobronchique
grâce à l’action de neuropeptides qui peuvent être classés en
deux catégories :
– le VIP (vasoactive intestinal peptide) et ses dérivés sont des
neurotransmetteurs de la composante inhibitrice du NANC par
action sur un récepteur membranaire musculaire spécifique ; le
monoxyde d’azote (NO) peut également intervenir dans cette
relaxation ;
– les tachykinines (substance P, neurokinines A et B) et le
CGRP (calcitonin gene-related peptide), libérés par les termi-
naisons nerveuses sensitives des fibres C bronchiques, sont les
neurotransmetteurs de la composante activatrice du NANC (8).
Médiateurs de l’inflammation
Le muscle lisse des voies aériennes possède également des
récepteurs membranaires spécifiques de médiateurs essentiel-
lement libérés par les cellules inflammatoires et participant à la
motricité bronchique.
L’histamine contracte le muscle bronchique par action sur le
récepteur de type H1 (9).
Les éicosanoïdes, dérivés du métabolisme de l’acide arachido-
nique, sont formés à partir des phospholipides membranaires
par action de la phospholipase A2. Les prostaglandines (PG) et
les leucotriènes (LT) possèdent des récepteurs spécifiques sur
la membrane du muscle bronchique (10). PGD2, PGF2αet
thromboxane A2 (TxA2) contractent la bronche humaine par
un même récepteur (récepteur TP), alors que PGE2 et PGI2
seraient relaxantes. Les éicosanoïdes pourraient participer à la
contraction bronchique induite par plusieurs substances,
notamment lors de la stimulation des récepteurs B2 et NK1
par, respectivement, la bradykinine et la substance P (11, 12).
Pour les leucotriènes, le muscle lisse bronchique humain se
contracterait davantage sous l’action de LTD4 que de LTC4 ou
de LTE4 en stimulant les récepteurs cysLT1.
Des médiateurs paracrines, notamment produits par les cellules
de l’épithélium bronchique, peuvent agir directement sur des
récepteurs spécifiques présents sur la membrane du muscle
bronchique. C’est le cas de l’endothéline (13). L’existence,
dans les bronches humaines, d’un facteur relaxant, dérivé de
l’épithélium (EpDRF) et pouvant agir sur un récepteur muscu-
laire spécifique, ainsi que sa nature restent encore controver-
sées. Le NO, qui pourrait, par analogie avec un des facteurs
relaxants dérivés de l’endothélium (EDRF), être l’EpDRF, a
des effets relaxants modestes sur le muscle lisse des voies
aériennes. Le NO n’agit pas sur un récepteur membranaire,
mais directement sur une enzyme cytoplasmique, la guanylate
cyclase.
Il n’est pas sûr que d’autres médiateurs inflammatoires comme
l’adénosine, le paf-acéther ou les radicaux superoxydes contrô-
lent directement le muscle bronchique par des récepteurs spé-
cifiques.
D’autres récepteurs, notamment aux cytokines, pourraient
moduler la fonction du muscle lisse sans avoir d’effet moteur
direct. Par exemple, il a été montré que l’interleukine 1ß aug-
mente l’expression des protéines Gi (inhibitrices de l’adénylate
cyclase) (14).
TRANSDUCTION MEMBRANAIRE : COUPLAGE EXCITATION-
CONTRACTION
Dans le muscle lisse des voies aériennes, le second messager
“activateur” est l’ion calcium. Les seconds messagers “inhibi-
teurs” sont les nucléotides cycliques AMPc et GMPc.
Récepteurs couplés aux protéines G
L’essentiel des récepteurs décrits ci-dessus appartient à la
famille des récepteurs membranaires couplés aux protéines G
(15). Ces protéines sont des hétérotrimères formées des sous-
unités α,β et γ. La liaison du messager extracellulaire à son
récepteur entraîne la substitution du GDP lié à la sous-unité α
de la protéine G par du GTP. La protéine G se dissocie alors en
deux sous-unités αet βγ. La sous-unité α-GTP active l’effec-
teur approprié, qui peut être une enzyme ou un canal ionique.
Tous les membres de cette famille des récepteurs couplés aux
protéines G ont une structure commune. Ils présentent un seg-
ment N terminal extracellulaire d’environ 30 acides aminés
(AA), sept domaines transmembranaires (TM1 à TM7) d’envi-
ron 25 AA chacun, reliés par une alternance de trois boucles
intracellulaires (i1 à i3), trois boucles extracellulaires (e1 à e3)
et une longue extrémité C terminale d’environ 80 AA. C’est la
nature de la protéine G couplée au récepteur qui détermine son
effet. Les récepteurs, dont l’activation entraîne une contraction
(par exemple H1, M3), sont en général couplés à une protéine G
de type Gq-11, dont la sous-unité αentraîne l’activation
d’une enzyme, la phospholipase C (PLC), impliquée dans la
signalisation calcique. D’autres récepteurs (par exemple, β2-
adrénergique ou M2) sont couplés à d’autres protéines G, res-
pectivement les sous-types Gs et Gi, qui augmentent ou dimi-
nuent la concentration d’AMPc par stimulation ou inhibition
de l’adénylyl-cyclase.
Signalisation calcique et contraction
L’augmentation de la concentration calcique cytoplasmique
peut être en rapport avec un influx de calcium extracellulaire
au travers des canaux calciques potentiel-dépendants. Cette
voie, peu utilisée par le muscle lisse des voies aériennes,
dépend du potentiel de membrane et donc de l’activité de nom-
7
La Lettre du Pneumologue - n° 2 - décembre 1998

breux canaux ioniques. L’influx de calcium extracellulaire
peut également s’établir au travers d’une autre catégorie de
canaux calciques, dont l’ouverture est commandée par l’inter-
action agoniste-récepteur. Il s’agit des ROC (Receptor-Opera-
ted-Channel). Enfin, et surtout, l’augmentation de la concen-
tration calcique cytoplasmique peut être liée à une libération
de calcium à partir des compartiments intracellulaires (réticu-
lum sarcoplasmique). L’IP3 formé grâce à l’action de la PLC
est le second messager qui libère le calcium intracellulaire
stocké dans les organites intracellulaires. L’influx de calcium
contribue à remplir de nouveau les compartiments intracellu-
laires.
Signalisation par les nucléotides cycliques et relaxation
La relaxation bronchique peut être considérée comme, d’une
part, l’antagonisme des mécanismes mis en jeu lors de la
contraction et, d’autre part, la mise en jeu de mécanismes spéci-
fiques à la relaxation. Les seconds messagers AMPc et GMPc,
impliqués dans la relaxation bronchique, interagissent de ma-
nière complexe avec le second messager Ca2+. Ces nucléotides
cycliques diminuent ou limitent l’augmentation de [Ca2+]i par
l’intermédiaire de protéine kinase AMPc-dépendante (PKA) et
GMPc-dépendante (PKG) (7). Ils interagissent en outre directe-
ment avec le fonctionnement des éléments contractiles.
Fonctionnement des éléments contractiles
Les protéines qui constituent la machinerie contractile du
muscle bronchique sont les mêmes que celles des autres
muscles lisses. La myosine (filament épais) est constituée de
deux chaînes lourdes (200-205 kDa) et quatre chaînes légères.
L’actine constitue un filament fin composé de deux polymères
de 42 kDa enroulé en hélice autour d’une autre protéine, la tro-
pomyosine. Deux autres protéines qui pourraient jouer un rôle
dans la contraction sont associées au filament fin, la caldesmone
et la calponine.
La phosphorylation réversible de la myosine constitue une
étape contrôlée préliminaire au développement de son activité
ATPasique en présence d’actine. Les chaînes légères de la
myosine sont phosphorylées grâce à une enzyme, la kinase des
chaînes légères de la myosine (MLCK). L’activité de cette
enzyme est contrôlée : activée par l’ion calcium après interac-
tion avec la calciprotéine et la calmoduline, et inactivée par
phosphorylation grâce à une kinase AMPc-dépendante (16).
Ainsi, la contraction du muscle lisse des voies aériennes fait
intervenir : l’augmentation de [Ca2+]i ; la liaison [Ca2+]i-calmo-
duline ; l’activation de la MLCK ; la phosphorylation des
chaînes légères de la myosine ; l’hydrolyse de l’ATP par
l’ATPase de la myosine phosphorylée combinée à l’actine ;
l’établissement de ponts actine-myosine qui s’attachent et se
détachent rapidement (“normally cycling cross bridges”),
entraînant le raccourcissement du muscle et la mise en tension.
La déphosphorylation de la myosine dépend d’une phospha-
tase des chaînes légères (MLCP).
Les mécanismes qui rendent compte de la contraction mainte-
nue sont encore controversés. Une hypothèse fait intervenir la
constitution de ponts d’actomyosine différents de ceux à trans-
lation cyclique (“normally cycling cross bridges”). Ces ponts
pourraient être attachés en permanence, “verrouillés” (“latch
bridges”), et ne nécessiteraient pas une phosphorylation préa-
lable de la myosine.
D’autres hypothèses sont proposées pour expliquer la contrac-
tion maintenue. Plusieurs faits expérimentaux suggèrent que la
protéine kinase C (PKC) pourrait être une enzyme intracellu-
laire fondamentale pour le maintien de la force.
Enfin, une hypothèse suggère que les protéines associées au
filament fin, caldesmone et calponine, pourraient être phos-
phorylées et agiraient sur la composante maintenue de la
contraction, peut-être en modifiant la sensibilité au Ca2+ de
l’appareil contractile. Ce concept de la modulation de sensibilité
au calcium de l’appareil contractile est d’actualité en physiolo-
gie et en pharmacologie cellulaire du muscle lisse bronchique.
Sa dysfonction pourrait participer à l’hyperréactivité non spé-
cifique observée dans l’asthme.
FONCTIONS DE SYNTHÈSE DU MUSCLE LISSE
BRONCHIQUE
La synthèse de médiateurs de l’inflammation par le muscle
lisse bronchique a été récemment mise en évidence (tableau I).
Ainsi, les cellules musculaires lisses bronchiques participent
au processus de l’inflammation bronchique. Cette fonction de
synthèse de la cellule musculaire lisse a été démontrée grâce
au développement des techniques de culture cellulaire.
Synthèse de cytokines et de chémokines
Il a été montré jusqu’à maintenant que le muscle lisse est sus-
ceptible de synthétiser les interleukines 1ß, 6 et 8, le facteur de
croissance GM-CSF, et les chémokines RANTES (regulated
upon activation normal T cell expressed and secreted) ou
MISE AU POINT
8
La Lettre du Pneumologue - n° 2 - décembre 1998
Médiateur synthétisé Facteur favorisant la synthèse du médiateur
IL1ß LPS
IL6 IL1ß, interféron γ
IL8 IL1ß, interféron γ, TNFα
GM-CSF IL1ß, interféron γ, ± TNFα
MCP-1 IL1ß, TNFα
RANTES IL1ß
IGF-2
IGFBP-2
ICAM-1 Interféron γ, TNFα
VCAM-1 TNFα
PDGF IL1ß
PGF2a, TxA2, PGE2, PGI2 Bradykinine, substance P, IL 1ß, interféron γ
NO LPS
Tableau I. Médiateurs synthétisés par les cellules musculaires lisse
GM-CSF : granulocyte-macrophage colony stimulating factor ;
ICAM : intracellular adhesion molecule ; IGF: insulin-like
growth factor ; IGFBP : IGF binding protein ; IL : interleukine ;
LPS : lipopolysaccharide ; PDGF : platelet-derived growth
factor ; PG : prostaglandine ; RANTES : regulated upon
activation normal T-cell expressed and secreted ; TNF
α
: tumor
necrosis factor
α
; Tx : thromboxane ; VCAM : vascular cell-
adhesion molecule.

MCP-1 (monocyte chemotactic protein) (17). Les interleukines
sont produites par le muscle lisse en culture incubé avec, par
exemple, du sérum de patient asthmatique, de l’interleukine
1ß, du TNFαet/ou de l’interféron γ. L’interleukine 1ß est une
des principales cytokines pro-inflammatoires. Elle augmente la
synthèse de nombreuses molécules : cytokines, molécules
d’adhésion, enzymes comme la cyclo-oxygénase 2, récepteurs
comme le récepteur B2 de la bradykinine. L’interleukine 6
induit non seulement la maturation des lymphocytes B et la
synthèse d’immunoglobulines, mais aussi l’activation et la dif-
férenciation des lymphocytes T. L’interleukine 8 active les
lymphocytes T et attire les neutrophiles. Le GM-CSF active et
augmente la survie des polynucléaires neutrophiles et éosino-
philes. RANTES a un rôle attractant sur les éosinophiles, les
neutrophiles et, comme MCP-1, sur les monocytes.
La part de la production de cytokines revenant au muscle lisse
sur l’ensemble des cytokines synthétisées dans l’asthme est
difficile à mesurer. Cependant, les études rapportées ci-dessus
suggèrent que le muscle lisse bronchique est susceptible de
potentialiser la réponse inflammatoire des cytokines pro-
inflammatoires comme l’interleukine 1ß, le TNFαou l’interfé-
ron γ.
Synthèse de facteurs de croissance du muscle lisse
Le remodelage des voies aériennes est un des éléments impor-
tants de l’obstruction dans l’asthme. Le muscle lisse est sus-
ceptible de subir des changements structuraux comprenant une
hyperplasie et une hypertrophie en réponse à des médiateurs de
l’inflammation comme l’histamine, l’endothéline, la tryptase
libérée par les mastocytes, des cytokines, des facteurs de crois-
sance, notamment le facteur de croissance dérivé des pla-
quettes (PDGF), le facteur de croissance épidermique (EGF) et
les facteurs de croissance “insuline-like” (IGF-1 et IGF-2).
Certains de ces facteurs de croissance, comme l’IGF-2 ou le
PDGF, peuvent être synthétisés par le muscle lisse lui-même
potentialisant ainsi sa propre croissance (18). La synthèse de
facteurs de croissance, notamment par le muscle lisse, est une
des cibles potentielles de thérapeutiques visant à réduire
l’hypertrophie et l’hyperplasie du muscle lisse bronchique.
Expression de molécules d’adhésion
Les cellules inflammatoires sont attirées dans la paroi bron-
chique par des facteurs chémotactiques produits par les cel-
lules épithéliales, les mastocytes, les polynucléaires éosino-
philes et, comme nous l’avons vu précédemment, les cellules
musculaires lisses. Celles-ci expriment à leur surface des
molécules d’adhésion spécifiques des lymphocytes T, le CD44
et l’ICAM-1 (intracellular adhesion molecule 1). L’expression
de ces molécules, induite par l’interféron γou le TNFα, permet
la fixation des lymphocytes T sur les cellules musculaires
lisses lors de l’inflammation (19). L’interaction entre lympho-
cyte et muscle lisse bronchique pourrait être un facteur déter-
minant d’entretien de l’inflammation, de l’hyperréactivité
bronchique et de la prolifération du muscle lisse. Il a été mon-
tré, par exemple, que les lymphocytes T pouvaient augmenter
la synthèse d’ADN par les cellules musculaires lisses.
Synthèse de prostaglandines
L’interleukine 1ß induit l’expression d’une cyclo-oxygénase de
type 2 (COX 2) dans les cellules musculaire lisses et, par
conséquent, la formation de prostaglandines relaxantes PGE2 et
PGI2, mais aussi contractantes PGF2α, PGD2 et TxA2 (20). La
PGE2 pourrait avoir un rôle important, mais qui reste à définir.
Elle est susceptible d’avoir un effet anti-inflammatoire car elle
inhibe la dégranulation des mastocytes, l’activation des éosino-
philes et la production d’IgE par les lymphocytes. Inhalée, elle
protège vis-à-vis de plusieurs agents bronchoconstricteurs.
Cependant, en augmentant la concentration d’AMPc intracellu-
laire, elle pourrait participer à la désensibilisation hétérologue
des récepteurs ß2-adrénergiques, induite par des médiateurs de
l’inflammation comme l’interleukine 1ß (21).
CONCLUSION
La mise en évidence de nouvelles propriétés du muscle lisse
bronchique le positionne comme un acteur essentiel de
l’hyperréactivité bronchique caractéristique de l’asthme. On le
savait capable de se contracter et de réduire ainsi le calibre
bronchique. Les mécanismes de cette contraction sont de
mieux en mieux connus et permettent d’envisager de nouvelles
voies de recherche thérapeutique. Par ailleurs, il est sujet à un
remodelage conduisant à une hypertrophie et à une hyperplasie
lors de l’asthme chronique, participant à l’obstruction et à
l’hyperréactivité bronchiques. Les mécanismes de ce remodelage
commencent à être démembrés et pourraient également consti-
tuer des cibles pharmacologiques. Enfin, il a été récemment
démontré que le muscle lisse peut participer activement à la
réaction inflammatoire. Cette cellule contractile peut mainte-
nant être considérée comme une cellule inflammatoire à part
entière. La découverte des mécanismes impliqués dans la régu-
lation des fonctions de synthèse du muscle lisse pourrait four-
nir de nouvelles cibles thérapeutiques dans l’asthme. ■
Références bibliographiques
1. Gabella G. Innervation of airway smooth muscle : fine structure. Ann Rev
Physiol 1987 ; 49 : 583-94.
2. Roffel A.F., Elzinga C.R., Zaagsma J. Muscarinic M3 receptors mediate
contraction of human central and peripheral airway smooth muscle. Pulm
Pharmacol 1990 ; 3 : 47-51.
3. Roux E., Molimard M., Savineau J.P., Marthan R. Muscarinic stimulation of
airway smooth muscle cells. Gen Pharmacol 1998 ; 31 : 349-56.
4. Fernandes L.B., Fryer A.D., Hirshman C.A. M2 muscarinic receptors inhibit
isoproterenol-induced relaxation of canine airway smooth muscle. J Pharmacol
Exp Ther 1992 ; 262 : 119-26.
5. Zhang Y., Molimard M., Advenier C. Influence of methocramine on the
acetylcholine-isoprenaline functional antagonism in the guinea-pig trachea.
Fund Clin Pharmacol 1996 ; 10 : 436-41.
6. Laitinen A., Partanen M., Hervonen A., Laitinen L.A. Electron microscopic
study on the innervation of the human lower respiratory tract : evidence of
adrenergic nerves. Eur J Respir Dis 1985 ; 67 : 209-15.
7. Devillier P., Molimard M., Bessard G., Advenier C. Aspects actuels des
récepteurs ß2-adrénergiques et des ß2-stimulants dans l’asthme. Rev Fr Allerg
1996 ; 36 : 269-81.
8. Barnes P.J. Neuropeptides in the lung : localization, function, and
pathophysiologic implications. J Allergy Clin Immunol 1987 ; 79 : 285-95.
9
La Lettre du Pneumologue - n° 2 - décembre 1998

MISE AU POINT
10
La Lettre du Pneumologue - n° 2 - décembre 1998
9. Marthan R., Crevel H., Guenard H., Savineau J.P. Responsiveness to
histamine in human sensitized airway smooth muscle. Respir Physiol 1992 ; 90 :
239-50.
10. Coleman R.A., Smith W.L., Narumiya S. International Union of
Pharmacology classification of prostanoid receptors : properties, distribution,
and structure of the receptors and their subtypes. Pharmacol Rev 1994 ; 46 :
205-29.
11. Molimard M., Martin C.A.E., Naline E., Hirsch A., Advenier C. Role of
thromboxane A2 in bradykinin-induced human isolated small bronchi
contraction. Eur J Pharmacol 1995 ; 278 : 49-54.
12. Naline E., Molimard M., Regoli D., Emonds-Alt X., Bellamy J.F., Advenier
C. Evidence for functional tachykinin NK1 receptors on human isolated small
bronchi. Am J Physiol 1996 ; 271 : L763-7.
13. McKay K.O., Black J.L., Armour C.L. The mechanism of action of
endothelin in human lung. Br J Pharmacol 1991 ; 102 : 422-8.
14. Hakonarson H., Herrick D.J., Serrano P.G., Grunstein M.M. Mechanism of
cytokine-induced modulation of beta-adrenoceptor responsiveness in airway
smooth muscle. J Clin Invest 1996 ; 97 : 2593-600.
15. Gilman A.G. G proteins : transducers of receptor-generated signals. Ann
Rev Biochem 1987 ; 56 : 615-49.
16. Savineau J.P., Marthan R. Activation properties of chemically skinned
fibres from human isolated bronchial smooth muscle. J Physiol (Lond) 1994 ;
474 : 433-8.
17. Johnson S.R., Knox A.J. Synthetic functions of airway smooth muscle in
asthma. Trends Pharmacol Sci 1997 ; 18 : 288-92.
18. Noveral J.P., Bhala A., Hintz R.L., Grunstein M.M., Cohen P. Insulin-like
growth factor axis in airway smooth muscle cells. Am J Physiol 1994 ; 267 :
L761-5.
19. Lazaar A.L., Albelda S.M., Pilewski J.M., Brennan B., Pure E., Panettieri
R.A. Jr. T lymphocytes adhere to airway smooth muscle cells via integrins and
CD44 and induce smooth muscle cell DNA synthesis. J Exp Med 1994 ; 180 :
807-16.
20. Belvisi M.G., Saunders M.A., Haddad el B., Hirst S.J., Yacoub M.H.,
Barnes P.J., Mitchell J.A. Induction of cyclo-oxygenase-2 by cytokines in human
cultured airway smooth muscle cells : novel inflammatory role of this cell type.
Br J Pharmacol 1997 ; 120 : 910-6.
21. Shore S.A., Laporte J. , Hall I.P., Hardy E., Panettieri R.A. Effect of IL1ß
on responses of cultured human airway smooth muscle cells to bronchodilatator
agonists. Am J Respir Cell Mol Biol 1997 ; 16 : 702-12.
1
/
5
100%