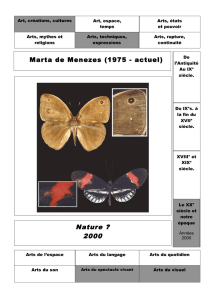
Propulsion biomimétique de structures élastiques

Propulsion biomim´etique de structures ´elastiques
Sophie Ramananarivo
To cite this version:
Sophie Ramananarivo. Propulsion biomim´etique de structures ´elastiques. M´ecanique des fluides
[physics.class-ph]. Universit´e Paris-Diderot - Paris VII, 2014. Fran¸cais. <pastel-00955323>
HAL Id: pastel-00955323
https://pastel.archives-ouvertes.fr/pastel-00955323
Submitted on 4 Mar 2014
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-
entific research documents, whether they are pub-
lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destin´ee au d´epˆot et `a la diffusion de documents
scientifiques de niveau recherche, publi´es ou non,
´emanant des ´etablissements d’enseignement et de
recherche fran¸cais ou ´etrangers, des laboratoires
publics ou priv´es.

THÈSE
Présentée à
UNIVERSITÉ PARIS DIDEROT
École doctorale :
MATIÈRE CONDENSÉE ET INTERFACES
(ED 518)
Par SOPHIE RAMANANARIVO
Pour obtenir le grade de DOCTEUR
Spécialité : MÉCANIQUE DES FLUIDES
PROPULSION BIOMIMÉTIQUE DE STRUCTURES ÉLASTIQUES
Thèse dirigée par :
BENJAMIN THIRIA ET RAMIRO GODOY-DIANA
Soutenue le : 10 JANVIER 2014
Devant la commission d’examen formée de :
M. Christophe Clanet Rapporteur
M. Christophe Eloy Rapporteur
M. Yves Couder Président
M. Emmanuel de Langre Examinateur
M. Jean-Marc Di Meglio Examinateur
M. Ramiro Godoy-Diana Directeur de Thèse
M. Benjamin Thiria Directeur de Thèse


TABLE DES MATIÈRES
Table des matières i
Remerciements 1
Résumé 3
1 Introduction 5
2 Interactions entre une structure élastique et le fluide environnant 23
2.1 Déformation d’un corps élastique . . . . . . . . . . . . . . . . . . . . 23
2.1.1 Elasticité . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
2.1.2 Poutre en flexion . . . . . . . . . . . . . . . . . . . . . . . . . . 26
2.1.3 Modes en milieu fini . . . . . . . . . . . . . . . . . . . . . . . . 29
2.2 Interactions fluide/structure . . . . . . . . . . . . . . . . . . . . . . . . 32
2.2.1 Problème couplé fluide/solide . . . . . . . . . . . . . . . . . . 32
2.2.2 Couplage fort : structure élancée en interaction avec un écou-
lement axial . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
2.3 Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
3 Vol battu 41
3.1 Modèle mécanique d’insecte auto-propulsé . . . . . . . . . . . . . . 41
3.1.1 Dispositif expérimental . . . . . . . . . . . . . . . . . . . . . . 41
3.1.2 Ailes de l’insecte . . . . . . . . . . . . . . . . . . . . . . . . . . 42
3.1.3 Paramètres de contrôle . . . . . . . . . . . . . . . . . . . . . . 45
3.1.4 Grandeurs mesurées . . . . . . . . . . . . . . . . . . . . . . . . 45
3.2 Résultats expérimentaux . . . . . . . . . . . . . . . . . . . . . . . . . . 48
3.2.1 Analyse dimensionelle : inertie, élasticité et forces fluides . . 48
3.2.2 Performances du système . . . . . . . . . . . . . . . . . . . . . 50
3.2.3 Cinématique des ailes . . . . . . . . . . . . . . . . . . . . . . . 53
3.2.4 Dynamique de l’amplitude et de la phase . . . . . . . . . . . 57
3.3 Modélisation théorique de l’aile flexible . . . . . . . . . . . . . . . . . 63
3.3.1 Modèle non-linéaire de poutre forcée . . . . . . . . . . . . . . 63
3.3.2 Résolution par échelles multiples . . . . . . . . . . . . . . . . 66
3.3.3 Mesure des coefficients de frottement de l’air ξet ξnl . . . . 69

ii Table des matières
3.3.4 Comparaison théorie/expérience . . . . . . . . . . . . . . . . 71
3.4 Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 72
3.4.1 Effets de résonance ? . . . . . . . . . . . . . . . . . . . . . . . . 73
3.4.2 Rôle de l’amortissement fluide . . . . . . . . . . . . . . . . . . 76
3.4.3 Optimum des performances . . . . . . . . . . . . . . . . . . . 76
3.5 Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 79
4 Nage Anguilliforme 81
4.1 Problématique . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81
4.2 Théorie des corps élancés de Lighthill . . . . . . . . . . . . . . . . . . 84
4.2.1 Force de propulsion . . . . . . . . . . . . . . . . . . . . . . . . 84
4.2.2 Point de vue énergétique . . . . . . . . . . . . . . . . . . . . . 87
4.3 Modèle mécanique de nageur anguilliforme . . . . . . . . . . . . . . 88
4.3.1 Nageurs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88
4.3.2 Dispositif expérimental . . . . . . . . . . . . . . . . . . . . . . 89
4.3.3 Paramètres de contrôle . . . . . . . . . . . . . . . . . . . . . . 92
4.3.4 Grandeurs mesurées . . . . . . . . . . . . . . . . . . . . . . . . 92
4.4 Résultats expérimentaux : performances et cinématique du nageur . 93
4.5 Réponse d’une poutre immergée à un forcage ponctuel . . . . . . . . 96
4.5.1 Modèle fluide/structure . . . . . . . . . . . . . . . . . . . . . . 96
4.5.2 Résolution numérique . . . . . . . . . . . . . . . . . . . . . . . 99
4.5.3 Comparaison prédictions théoriques - mesures expérimen-
tales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 103
4.5.4 Rôle prépondérant du frottement fluide quadratique . . . . 104
4.6 Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 107
4.6.1 Prédiction de la vitesse de nage, modèle de Lighthill . . . . . 107
4.6.2 Efficacité mécanique . . . . . . . . . . . . . . . . . . . . . . . . 109
4.7 Modèle de traînée globale . . . . . . . . . . . . . . . . . . . . . . . . . 112
4.8 Propagation d’ondes en milieu élastique fini . . . . . . . . . . . . . . 120
4.8.1 Equation dynamique d’une plaque forçée . . . . . . . . . . . 121
4.8.2 Expériences de vibration . . . . . . . . . . . . . . . . . . . . . 123
4.8.3 Influence de la dissipation sur la cinématique de la plaque . 124
4.8.4 Perspectives . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 129
4.9 Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 130
5 Conclusion et perspectives 133
Bibliographie 137
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
1
/
150
100%