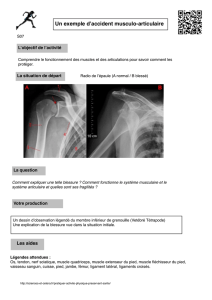
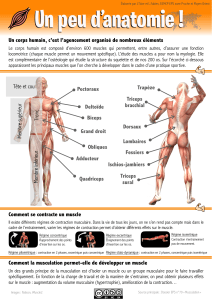
Muscle, Myographie


!"#
!!#
$
!$
$
#%&&$
#!'()*!+$
#',*()*+-
##')()*#+.
#$'))()*$+.
$ %).
$!')()*-+.
$/0
- 0
. %)**12
0 3&
0!3&!4567
03&43!

#
!
"
881)
1(+78*9:1
(;)+
<*7*6&7*
61)*=6&>
?77;7@
6=1;
,7;()
&!467+
!!
A1;)1;6;;
,;(;+=;
;&B18,1(+C;
=;7&, *1(
1+1;&1(:
D
+=7*
1(1+);1
7*B1;&*C*
DD
1
=*7&,($
DD
+1
)7;*7&,";
7;?);3
1;3A(7&+(1+@>3A7&,
;);>3A&1;;
;3>3;;7&>E,;
16@17&,
1;*?;1,;)('F!
#-+
/!4,;
("'F!#-+
=;?1*7*,
1,DD
;3=;111G7&
;

$
!
H*)1()
&4+;3
81' 35
7I
=6?7&)7
,(;#,-+
=6,1*
7?;1
?*;7;*
&)
#
%&&
#!'()*!+
5)7;*
;&&
=*
@
E
*4
4!#2
40.-
"4.2(JK$;),
+
>E(,;&+4!-#0
"4
=4
"6;);
B18,1*7&;17*
@&1:
D
1
;7*B1;&**
DD
1
=4
'6()!"+

-
5**1;&
**5&16LM,
E61I('F!#.+
/461I
("'F!#0-+
=6I6(+6
1()B)+=1?
11)1;,;
=E4
=E
DD
11
1?7;A
;&E?)
;*
DD
516
)11,
%1;&4
=;&;&
,;&7*)
;*;L9M*
;
#',*()*+
*,.2!
3!;**,*
616
32;***,!;1
*16
:**,&;E
6;;7=
*(N$$$+
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%