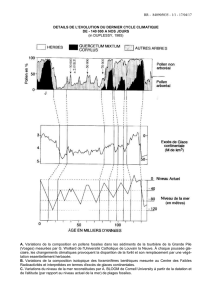
Les associations fossiles Les associations fossiles

1
Les associations fossiles
Paléobiocénose
Elts allochtones
Elts autochtones
Taphocénose
Les Lagerstättes
Les Ambres
Pièges sédimentaires
(e.g.Karst)
Paléo- écosystemes partiellement
conservés, considérations paléo-
écologiques possibles
Mélanges écologique et temporel,
analyses des données nécessaires
Les associations fossiles
Behrensmeyer et al., 2000 Paleobiology

2
Paléobiocénose: tous les fossiles présents sont considéré autochtones (cas
des gisements à formes sessiles en particulier). Cette condition est bien
entendu aussi vérifiée dans le cas des Lagerstattes. Néanmoins on peut le
vérifier si l'on observe :
)l'absence de trace de transport
)l'absence de désarticulation majeure du fossile (intégralité du squelette)
)Une représentation des différents stades de vie (surtout pour les organismes
sessiles, hors lagerstatte) par l’analyse statistique de caractères liés à la
croissance (taille, poids, etc..)
)Présence de témoins d’une activité biologiques (piste, coprolithes, etc…)
)Caractères sédimentaires de l'encaissant.
Les Lagerstättes
(fossil-lagerstätten)

3
Les Lagerstättes (fossil-lagerstätten)
* Ediacara (Australie -PréCambrien)
* Niryani (Hongrie- Cambrien)
* Schistes du Burgess (Canada- Cambrien)
* Gisement de Montceau-les-mines (France - Stephanien)
* Gisement de Bear Gulch (Carbonifère - Mississipien)
* Monte San Giorgio (Italie – Trias)
* Holzamden (Allemagne - Jurassique inférieur)
* La Voulte-sur-Rhône - ( France – Jurassique moyen)
* Solnhofen (Bavière - Jurassisque sup)
* Canjuers-Cerin (France - Jurassique supérieur)
* Monts du Liban (Santonien)
* Creil (Oise - Paleocene)
* Menat (Puy de dôme - Eocene)
* Green River (répartis sur 3 états des Etats-Unis- Eocene)
* Monte Bolca (Italie - Eocene)
* Gisement de Messel (Allemagne - Eocene)
* Rancho La Brea (Californie - Quaternaire)
Les Lagerstättes (fossil-lagerstätten)
Solnhofen -Tithonien

4
Les Lagerstättes (fossil-lagerstätten)
La Voulte sur Rhône (Ardèche) - Callovien
Les Lagerstättes (fossil-lagerstätten)
Messel - Eocène
Bactérie fossilisées sur la trace des tissus mous d’une
chauve sourie de MESSEL (Eocène)

5
Ambres
Différentes variétés de l'ambre opaque crétacé des Charentes, de couleur miel et un peu translucide ou brun laiteux plus ou moins
sombre ; b et c) microradiographies d'un morceau d'ambre opaque, par b) imagerie d'absorption et (c) contraste de phase, révélant,
à droite et à gauche, les silhouettes de deux insectes (ESFR, 2008)
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
1
/
45
100%