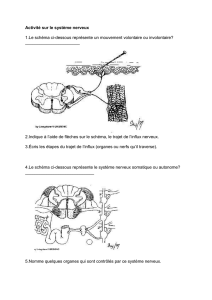
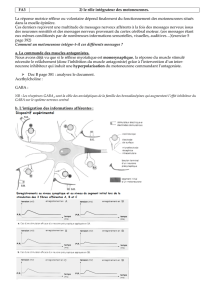
Le communication nerveuse

Partie 2 :
Le communication nerveuse

TP6
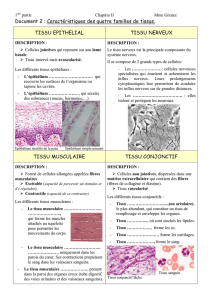
Chapitre 1 :
Le réflexe myotatique, une
réponse musculaire adaptée et
coordonnée.
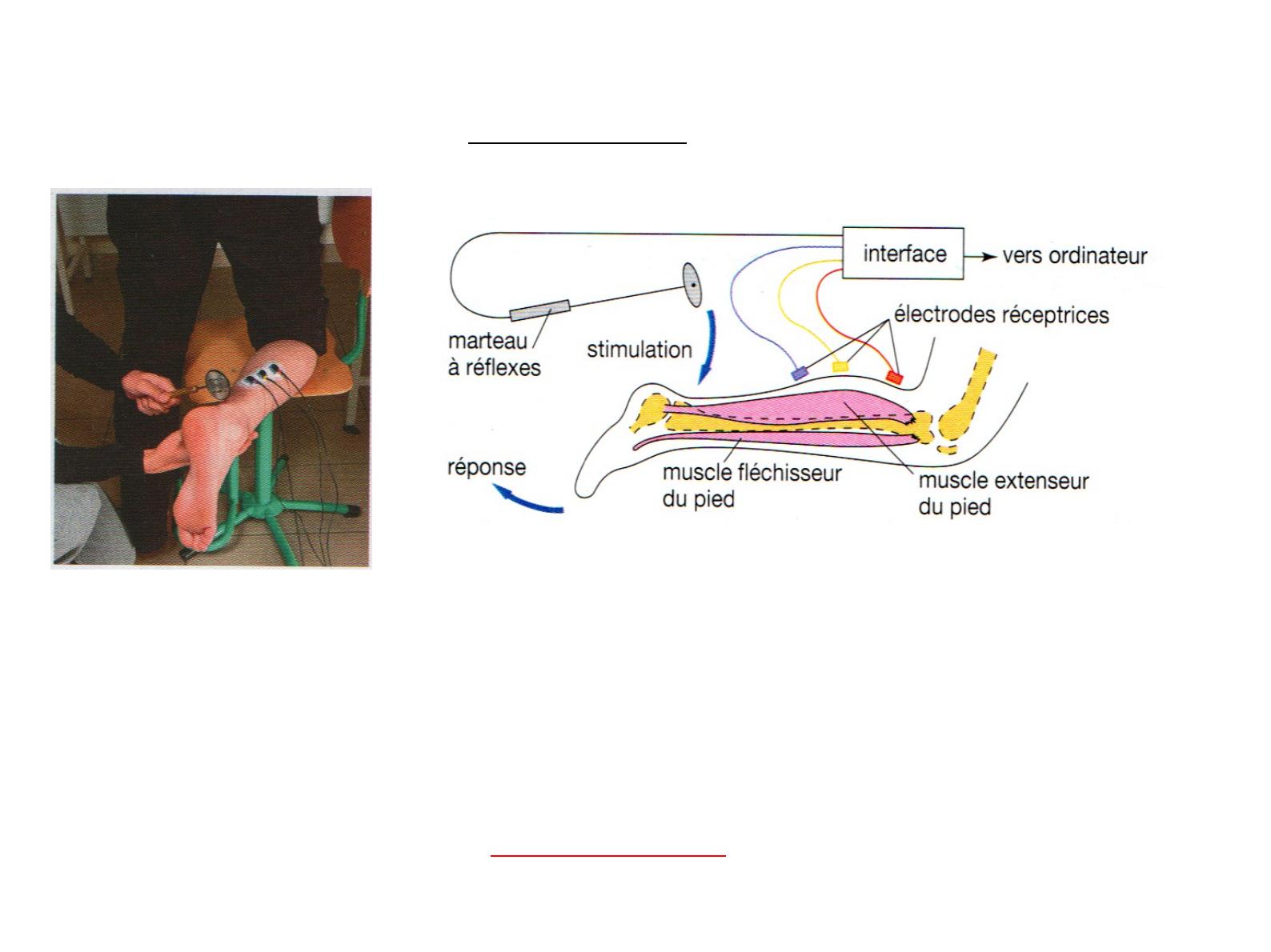
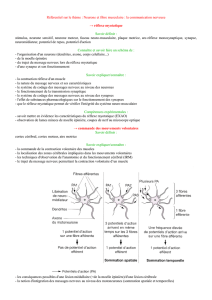
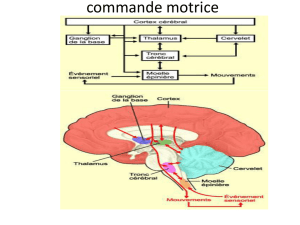
Le maintien de la posture nécessite un réajustement permanent et réflexe, de l'état de
contraction des muscles appelé tonus musculaire. Le réflexe myotatique est l’élément
principal de ce maintien.
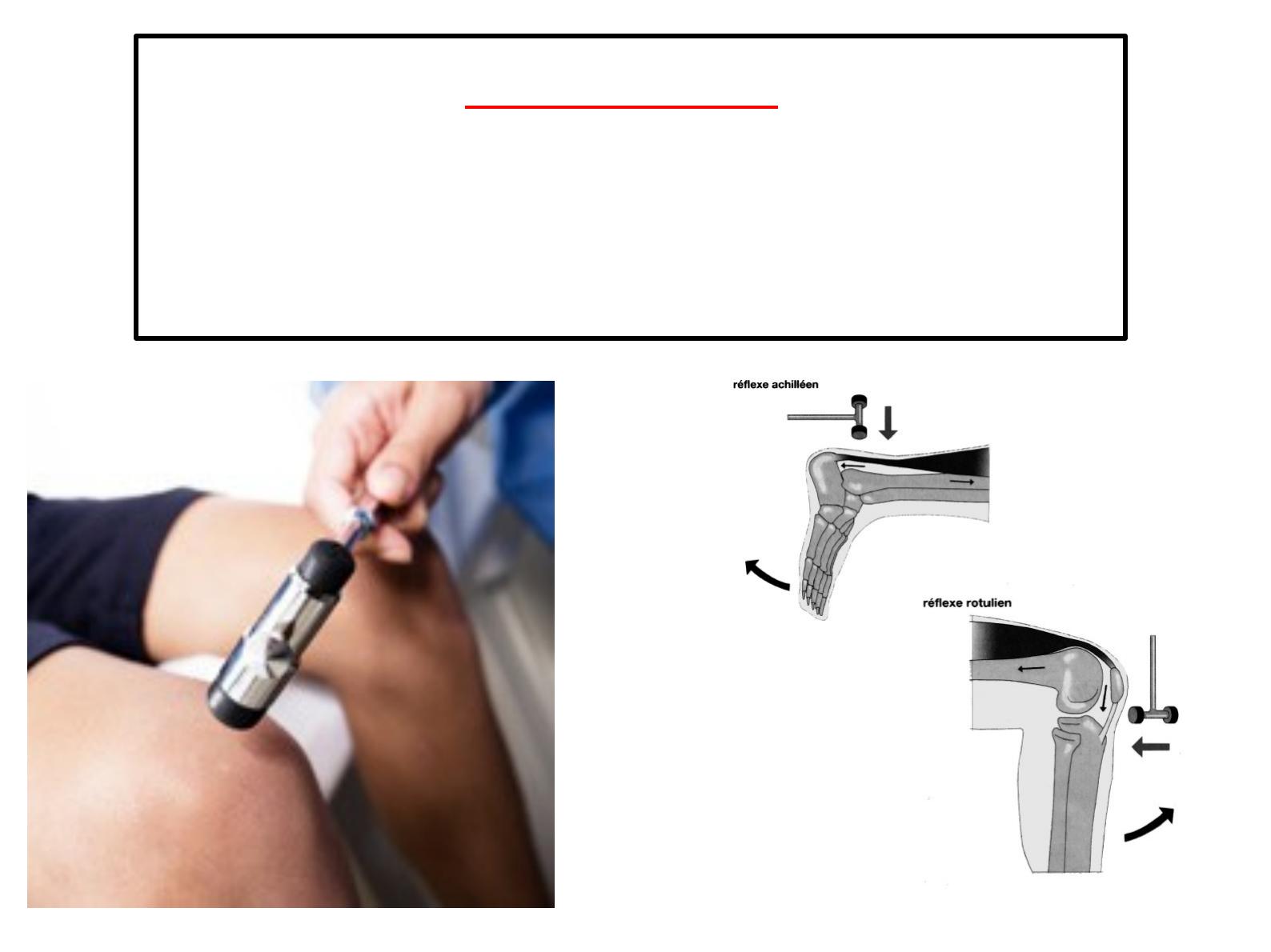
Un coup sec porté sur le tendon d’Achille à l’aide d’un « marteau à réflexes »
entraîne un mouvement involontaire d’extension du pied : on parle de réflexes
achilléen.
De même un coup sec porté au dessous de la rotule provoque immédiatement
l’extension de la jambe…
On parle dans tous ces cas de réflexe myotatique (contraction reflexe d'un muscle
en réponse à son propre étirement).
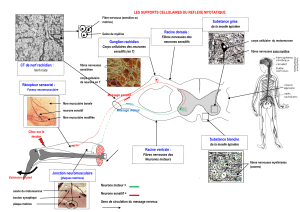
1. Principe et définition:
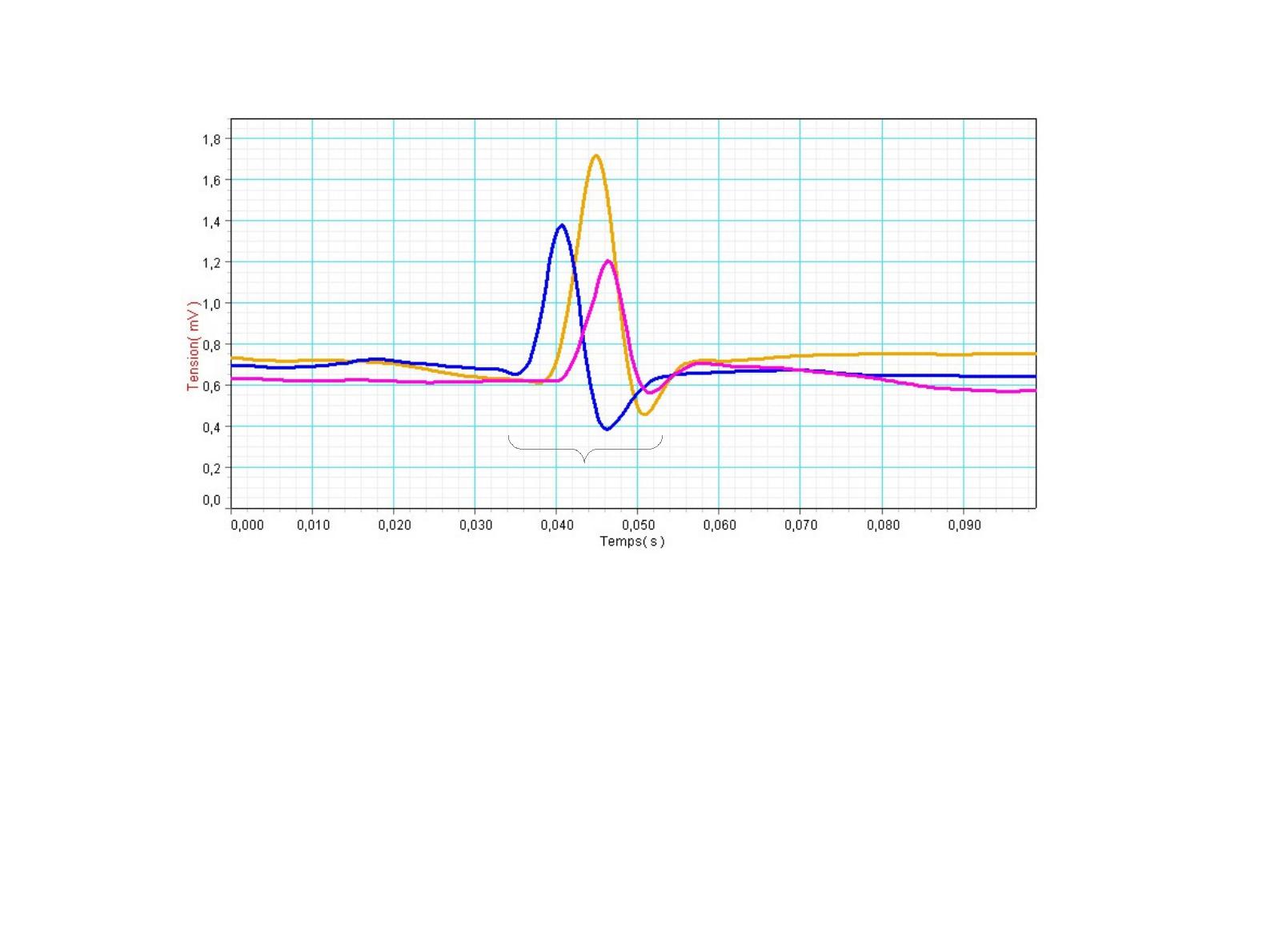
Contraction musculaire
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
1
/
68
100%