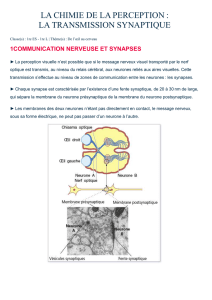
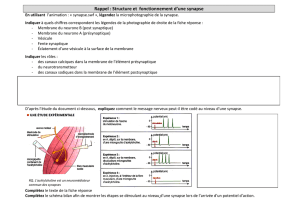
UV_101_BASES_DE_PHYSIOLOGIE_NEURO

1
BASES DE PHYSIOLOGIE
UV 101
SYSTEME NERVEUX
Neurones et synapses
P. PILARDEAU

2
SYSTEME NERVEUX
Le système nerveux est constitué de cellules spécialisées appelées neurones. Ces cellules sont
localisées essentiellement dans le cerveau, le cervelet, le tronc cérébral et la moelle épinière.
Les neurones entrent en communication entre eux (synapses) ou avec d’autres tissus
(musculaires, glandulaires).
Le système nerveux permet de capter des informations extérieures à l’organisme (visuelles,
auditives, sensitives, gustatives...), d’intégrer ces données (zones spécialisées du cerveau en fonction
de du stimulus) et de fournir une réponse la plus adaptée possible (contraction musculaire, sécrétion
glandulaire...).
I - ANATOMIE
I-I - LE NEURONE
Le neurone, ou cellule nerveuse, est formé d’un corps cellulaire (encore appelé soma),
comportant l’ensemble des organites trouvées dans les cellules (noyau, ribosome, mitochondries...) et
des prolongements donnant à la cellule un aspect étoilé. Ces prolongements sont de deux types :
Courts et nombreux, il s’agit des dendrites.
Long et unique, il s’agit de l’axone
Les dendrites ont pour fonction de recevoir des messages destinés aux neurones. L’axone
prend naissance dans le soma, il est long de 1 à 50 microns, son rôle est de transmettre le message
neuronal à d’autres cellules.
Le neurone
Dendrites
Axone
Soma
1.1.1 Le soma
Les neurones sont groupés dans le système nerveux central où ils forment des structures
nodulaires appelées noyaux gris centraux. Dans le système nerveux périphérique les corps
cellulaires forment les ganglions latérovertébraux (attention, il s’agit de structures nerveuses et non
des ganglions lymphatiques que l’on peut parfois percevoir sous la peau).
Le cytoplasme du neurone, comme c’est le cas pour toutes les cellules de l’organisme, est très
concentré en potassium (100 mEq/l), pauvre en chlore (20 mEq/l) et pauvre en sodium (20 mEq/l). Il
existe donc un déséquilibre important entre le milieu intracellulaire et le liquide extracellulaire très
concentré en sodium (140 mEq/l), et en chlore (110 mEq/l) et relativement faible en potassium (3
mEq/l).

3
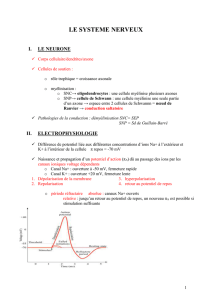
Ces caractéristiques sont à l’origine d’une différence de potentiel de 90 mV (gradient
électrique) et d’un gradient chimique (Na+/K+). Au repos, c’est à dire en dehors de toute stimulation,
le corps cellulaire est donc négatif (- 90 mV).
Intra cellulaire Extracellulaire
Na + 20 142
K + 100 4
Ca++ Traces 4
Cl - 20 100
HCO3- 8 27
Autres 152 23
anions (-)
Bilan
ionique 300 mEq 300 mEq
ou 300 mOsm ou 300 mOsm
Charge
électrique - 70 mV 0 mV
Sens des gradients :
Les différences chimiques et électriques entre l’intérieur et l’extérieur de la cellule sont à
l’origine de gradients transmembranaires concernant le potassium et le sodium.
Intracellulaire Extracellulaire
Gradient chimique Na Na
K K
Gradient électrique Na+
- 70 mV
K+

4
Le gradient chimique tend à faire entrer le sodium dans la cellule et à extruder le potassium. Le
gradient électrique attire vers l’intérieur de la cellule les particules chargées positivement
(sodium et potassium).
1.1.2 L’axone
L’axone, ou substance blanche (par opposition aux corps cellulaires qui apparaissent gris)
transmet le message du corps cellulaire à d’autres cellules (nerveuses ou non). Les axones sont
regroupés en faisceaux, constitutifs des nerfs.
L’axone, ou fibre nerveuse, peut être entouré d’une gaine phospholipidique appelée gaine de
myéline. Cette dernière se trouve interrompue au niveau de zones d’étranglement (étranglement de
Ranvier).
Cette gaine, électriquement isolante, est sécrétée par les cellules de Schwann qui entourent
l’axone. Le cylindraxe proprement dit est constitué de la même substance que le soma, il est parcouru
par un faisceau de neurofibrilles.
Myéline Nœud de Ranvier
Axone
Certaines fibres sont dépourvues de myéline, il s’agit des fibres amyéliniques, pratiquement
spécifiques des nerfs sympathiques.
1.1.3 - Membrane neuronale
La membrane des neurones comprend des zones spécifiques d’échange appelées canaux
ioniques. Ces canaux sont chargés d’assurer et de réguler les échanges entre l’intérieur et l’extérieur
de la cellule.
On distingue :
= Les canaux passifs
= Les canaux voltage-dépendant
= Les canaux ATP dépendant
Les canaux passifs :
Ces structures sont très perméables à l’ion K+ et relativement peu à l’ion Na+.

5
Extérieur du neurone
Na+ K+ Ca++
Membrane Membrane
K+
Intérieur
Canal à potassium
Le canal à K+ laisse sortir les ions potassium dans le milieu extérieur du fait du gradient de
concentration, tout en bloquant la pénétration des ions Na+ et Ca++, pourtant également chargés
positivement.
Au sein du canal existe quatre charges négatives ( ) qui interdisent toute entrée d’anion Cl-
isolé ou d’autres anions.
Pour sortir de la cellule, le potassium se lie aux charges négatives du canal (neutralisation
de la charge) et ce, de manière très spécifique (à ce niveau le Na+ ne peut entretenir de liaison forte
avec un anion). Cette double spécificité permet le passage du potassium à l’exclusion de celle du
sodium ou du calcium.
Au repos ces pores (ou ces canaux) sont ouverts. La perméabilité au potassium est très
importante.
Les canaux voltage-dépendant
Ces canaux sont activés par des stimuli électriques physiologiques.
Ces canaux permettent les passages de Na+ vers l’intérieur de la cellule et la sortie
simultanée de potassium, inversant ainsi la polarité de la membrane cellulaire.
On distingue :
Les canaux Na+ comprenant une double porte (vers l’intérieur et vers l’extérieur). Et
les canaux K+ ne comportant qu’une seule porte.
Les canaux sodium sont fermés au repos, la membrane est très perméable au
potassium et pratiquement imperméable au sodium.
 6
7
8
9
10
11
12
13
14
15
16
6
7
8
9
10
11
12
13
14
15
16
1
/
16
100%