fonctionnement du systeme nerveux

URL source du document
www.lucieberger.org/svt
Document
FONCTIONNEMENT DU SYSTEME
NERVEUX
Nous allons acquérir les notions de base sur la communication nerveuse, puis élargir la
compréhension des relations entre le phénotype et le génotype, pour étudier la part de
l'expérience individuelle et celle de l'hérédité dans le remodelage permanent des circuits
nerveux.
Eléments requis:
L'intégration des fonctions dans l'organisme
Thèmes traités et activités pédagogiques (durée: 6 semaines):
Les circuits neuroniques d'un réflexe
Les messages nerveux
Plasticité du système nerveux central
Thème 1: Les circuits neuroniques d'un réflexe
La réalisation d'un mouvement nécessite la contraction de muscles précis. Commandée par le
système nerveux, elle peut correspondre:
soit à un mouvement volontaire (activité spontanée),
soit à un mouvement automatique, involontaire en réaction à une stimulation (activité
réflexe).
1.1 LE REFLEXE MYOTATIQUE:
Un réflexe se définit comme une réponse d'origine nerveuse, indépendante de
la volonté, automatique, en réplique à une stimulation. Dans le cas d'un réflexe
myotatique, il s'agit de la contraction réflexe d'un muscle déclenchée par son
propre étirement. De tels réflexes interviennent dans le maintien de la posture
pour s'opposer aux effets de la gravité. Ils sont testés par le médecin (réflexe
rotulien, réflexe achilléen). Ce dernier, par exemple est déclenché par un choc
bref sur le tendon d'Achille et se manifeste par une extension du pied,
conséquence de la contraction du triceps sural du mollet.
1.1.1 Enregistrement du réflexe achilléen par ExAO
Principe de l'enregistrement: quand un muscle se contracte, il est le
siège d'une activité électrique. A l'aide d'électrodes réceptrices et d'un
amplificateur (module électrophysiologique), il est possible

d'enregistrer cette activité: c'est l'électromyographie. Les
tracés obtenus (électromyogrammes) traduisent ainsi
l'activité électrique musculaire.
Protocole: trois électrodes sont placées sur le muscle du mollet
(triceps sural = extenseur du pied) comme l'indique le dessin ci-
contre à droite.
Le réflexe est déclenché en frappant à l'aide du marteau qui est
connecté au dispositif d'ExAO (schéma ci-contre à gauche). Au
moment du choc sur le tendon d'Achille, un contact électrique
déclenche un signal qui détermine l'enregistrement des données fournies par
les électrodes et leur traitement par le biais du logiciel installé sur l'ordinateur.
En même temps, cela provoque la contraction réflexe du muscle extenseur du
pied.
Enregistrement:
Vous réaliserez cette expérience et vous imprimerez
l'enregistrement obtenu. En voici un exmplaire ci-dessous:
Repérez sur l'enregistrement le délai qui existe entre l'instant du choc et celui
de la réponse musculaire. Sachant que la contraction musculaire est
déclenchée par des messages nerveux en provenance de la moelle (et non
directement par l'étirement du tendon) déterminez le trajet du message nerveux
responsable de la réponse réflexe. Evaluez-le et donnez ainsi un ordre de
grandeur de la vitesse du message nerveux.
Si vous cherchez à modifier l'intensité du stimulus porté sur le tendon
d'Achille, que révèle l'intensité de la réponse réflexe?
1.1.2 Des éléments anatomiques et histologiques interviennent dans la mise
en oeuvre du réflexe achilléen. Nous les étudierons dans le cadre d'un T.P.
d'histologie, leur concours à la réalisation du réflexe sera justifiée juste après.
Ce sont:

des mécano-récepteurs, ici des fuseaux neuro-musculaires, stimulés
par l'étirement: éléments constitués d'une capsule fibreuse
contenant des fibres musculaires entourées des terminaisons de
fibres ( ces dernières sont dénommées Ia sur la figure que vous
découvrirez en cliquant sur l'icône ci-contre->). Ils sont placés en
parallèle des fibres musculaires. Les fuseaux neuro-musculaires
détectent la longueur des muscles et leur tension et transmettent cette
information par les fibres afférentes* Ia.
* on qualifie d'afférentes les fibres nerveuses qui partent des récepteurs
sensoriels, tels que les fuseaux neuromusculaires et qui arrivent au centre
nerveux comme la moelle épinière. Par opposition, on qualifie d'efférentes les
fibres nerveuses qui partent du centre nerveux pour se diriger vers les
effecteurs périphériques, tels que les muscles.
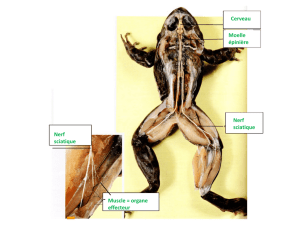
des fibres nerveuses sensitives, logées dans le nerf rachidien sciatique,
qui conduisent à la moelle les messages afférents ou centripètes,
des fibres nerveuses motrices du nerf sciatique, transmettant les
messages efférents ou centrifuges aux effecteurs "muscles".
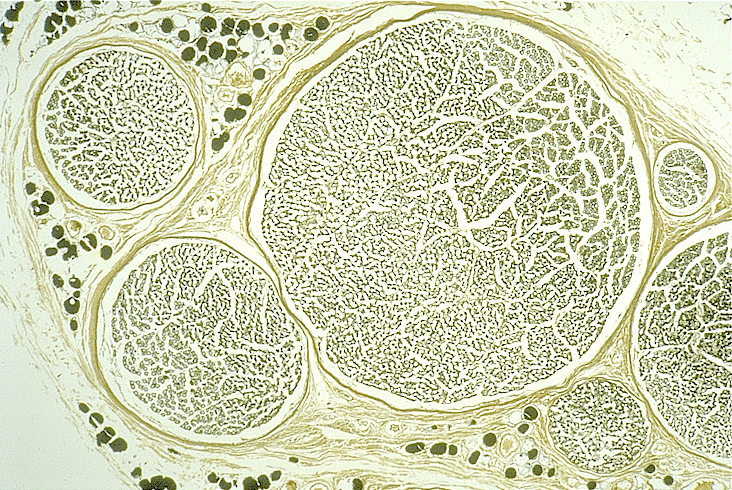
Vous observerez deux préparations de nerf rachidien du
commerce:
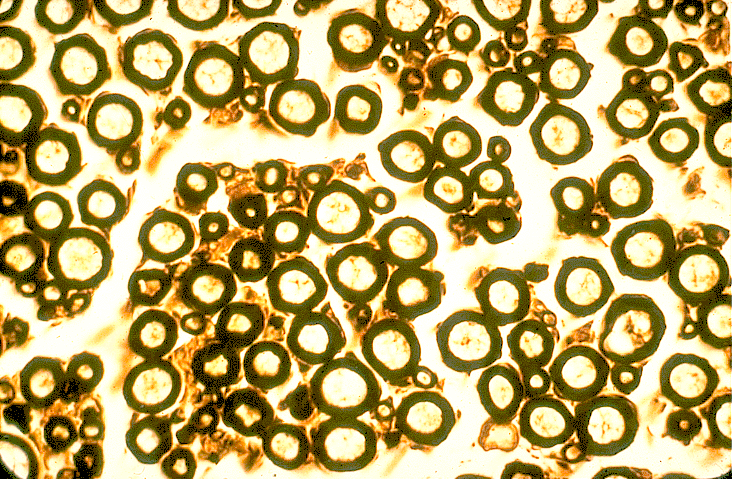
- une coupe transversale de nerf rachidien, tel que le sciatique qui innerve le
membre inférieur (ou postérieur chez les animaux),
- une préparation de nerf dilacéré qui vous montrera des fibres nerveuses en
long.
Le portail du Department of Biological Sciences - University of Delaware
(Dr. Roger C. Wagner and Dr. Fred E. Hossler) vous fournira aussi
d'excellentes vues microscopiques:
- coupe transversale de nerf périphérique traité à l'acide osmique (faible
grossissement),
- coupe transversale de fibres myélinisées dans un nerf périphérique traité à
l'acide osmique (fort grossissement),
- fibres dilacérées d'un nerf périphérique montrant les noeuds de Ranvier (fort
grossissement),
Vous réaliserez des dessins de vos observation de manière à relater la
structure d'un nerf et de ses fibres.
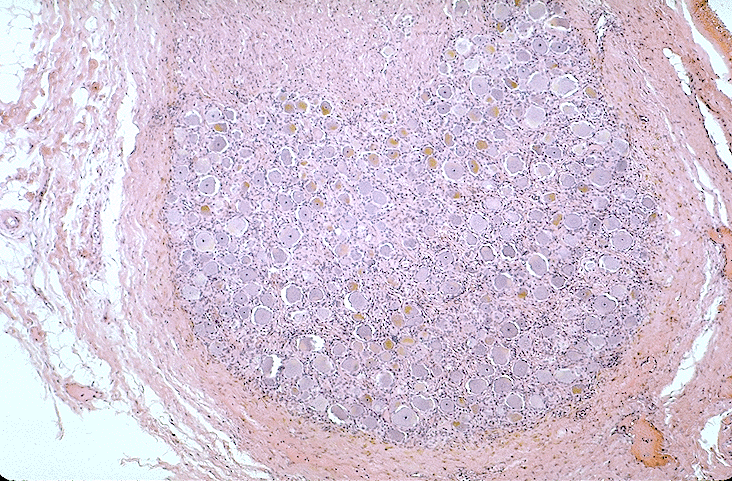
la moelle épinière lombaire, centre nerveux réflexe. Son organisation
anatomique et notamment ses relations avec chacun des nerfs
rachidiens pairs, auxquels elle est reliée par deux racines, l'une dorsale,
comportant un ganglion rachidien et l'autre ventrale, peut être observée
sur une coupe transversale observée à la loupe ou au très faible
grossissement du microscope.

Vous observerez au microscope une coupe transversale de
moelle épinière à différents grossissements.
Le portail du Département d'Histologie et d'embryologie de la faculté
Universitaire FUNDP de NAMUR, dans lequel vous sélectionnerez la rubrique
"Histologie générale - Tissus nerveux - Architecture schématique - La moelle
épinière", vous permettra d'être guidé dans votre étude.
les muscles eux-mêmes, dont les myofibres avec leur plaque motrice
individuelle, constituent les effecteurs. Il est ainsi possible de définir
ce qu'est une unité motrice: c'est l'ensemble constitué par un neurone
et ses ramifications, assurant la commande synchrone d'un certain
nombre de fibres musculaires. Le microscope électronique permet de
comprendre l'intimité de la jonction neuro-musculaire (synapse).
Tous ces éléments sont intégrés dans un circuit constituant l'arc réflexe, de
type médullaire ici, dont nous allons justifier de l'intervention.
1.2 LES CIRCUITS NERVEUX DU REFLEXE
1.2.1 Les expériences de dégénérescence wallérienne:
Ces expériences historiques ont été réalisées par Augustus Desiré WALLER en
1850.
Lorsque le corps cellulaire d'un neurone est détruit, la fibre nerveuse dégénère
totalement. Lorsque la fibre nerveuse est sectionnée, la partie distale, c'est à
dire la partie qui est séparée du corps cellulaire, dégénère. C'est le phénomène
de la dégénérescence wallérienne. Par contre la partie proximale, c'est à dire
celle qui est restée en rapport avec le corps cellulaire, peut régénérer.
Waller a réalisé chez l'animal des
sections de tronçons nerveux, notamment
celles représentées sur les schémas ci-contre
à droite. Donnez une interprétation aux
résultats de chacune d'elles dans le but de
préciser finalement où se trouvent les corps
cellulaires des neurones intervenant dans le
réflexe médullaire.
1.2.2 Les expériences de Bell-Magendie:

Charles BELL,
dès 1811,
entreprit de
définir le rôle
des racines
rachidiennes
dans la
conduction
nerveuse.
C'est François
MAGENDIE
en 1822 qui
précisa le rôle
des racines
antérieures et
postérieures
des nerfs
rachidiens, Charles BELL le confirma en 1824.
Une variante de ces expériences est présentée ci-contre à gauche: elles
consistent à opérer, après anesthésie des seuls centres supérieurs (cerveau)
chez un animal (animal spinal), des sections de racines rachidiennes, de les
stimuler et d'observer les effets de ces stimulations.
Analysez les résultats de ces expériences et tirez les conclusions qui
s'imposent.
1.2.3 Deux types de neurones sont mobilisés:
Il est à présent possible d'identifier les deux types de neurones intervenant
obligatoirement dans un réflexe:
- les neurones sensitifs afférents dont les influx centripètes cheminent des
récepteurs musculaires à la moelle en passant par les racines dorsales des nerfs
rachidiens: ils ont leurs péricaryons (corps cellulaires nucléés) dans les
ganglions rachidiens.
Là encore le portail du Département d'Histologie et
d'embryologie de la faculté Universitaire FUNDP de NAMUR, dans lequel vous
sélectionnerez la rubrique "Histologie générale - Tissus nerveux - Neurones -
§ Les cellules nerveuses ganglionnaires des ganglions rachidiens ou spinaux"
vous permettra de reconnaître les neurones ganglionnaires en T.
Vous en ferez un dessin d'observation, après avoir réalisé une observation sur
une préparation du commerce.
- les motoneurones efférents dont les influx centrifuges empruntent les
racines ventrales des nerfs rachidiens et parviennent aux muscles: leurs
péricaryons se situent dans les cornes antérieures de la moelle (neurones
multipolaires).
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
1
/
21
100%









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}