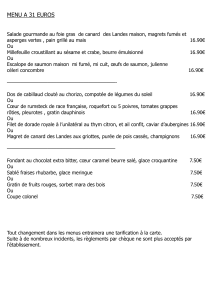
étude du portage d`agents pathogènes respiratoires chez

ANNEE 2005 THESE : 2005 – TOU 3 – 4044
ÉTUDE DU PORTAGE D’AGENTS PATHOGÈNES
RESPIRATOIRES
CHEZ LE CANARD MULARD
ET CONSÉQUENCES CLINIQUES
_________________
THESE
pour obtenir le grade de
DOCTEUR VETERINAIRE
DIPLOME D’ETAT
présentée et soutenue publiquement en 2005
devant l’Université Paul-Sabatier de Toulouse
par
Cyril, Georges, Emile ARMAND
Né, le 18 mai 1979 à ANNECY (Haute-Savoie)
___________
Directeur de thèse : Monsieur le Docteur Jean-Luc GUERIN
___________
JURY
PRESIDENT :
M. Henri DABERNAT
ASSESSEUR :
M. Jean-Luc GUERIN
M. Guy BODIN
Professeur à l’Université Paul-Sabatier de TOULOUSE
Maître de Conférences à l’Ecole Nationale Vétérinaire de TOULOUSE
Professeur à l’Ecole Nationale Vétérinaire de TOULOUSE

Toulouse, 2005
-
NOM : ARMAND PRENOM : CYRIL
TITRE : ETUDE DU PORTAGE D’AGENTS PATHOGENES RESPIRATOIRES CHEZ LE
CANARD MULARD ET CONSEQUENCES CLINIQUES
RESUME :
Les syndromes respiratoires constituent une dominante pathologique de l’élevage du canard
mulard. Plusieurs agents bactériens (Pasteurella multocida, Riemerella anatipestifer,
Escherichia coli, Ornithobacterium rhinotracheale, Mycoplasma sp., Chlamydophila
psittaci), fongiques (Aspergillus fumigatus), ou viraux (Metapneumovirus du canard) peuvent
être impliqués dans ces syndromes. Cependant, ces germes peuvent également faire l’objet
d’un portage sain et le rôle respectif de ces agents et des facteurs de risque technique reste à
préciser. L’objectif de ce travail est d’évaluer le portage de ces agents sur les canards mulards
en élevage et de mettre en relation ce portage avec le statut clinique des animaux. Une étude
cas témoin a été réalisée entre février et juin 2004 dans le Sud-Ouest. Dix huit lots de canards
mulards ont été prélevés et analysés. Si le portage d’agents pathogènes respiratoires apparaît
important et polymorphe, il n’a pu être établi de relation causale statistiquement significative
entre ce portage et le statut clinique des animaux.
MOTS-CLES : MALADIE RESPIRATOIRE / CANARD MULARD / CHLAMYDOPHILA
/ RIEMERELLA / PASTEURELLA / ORNITHOBACTERIUM / ESCHERICHIA COLI /
MYCOPLASMA / ASPERGILLUS / METAPNEUMOVIRUS
___________________________________________________________________________
ENGLISH TITLE : A SURVEY ABOUT THE CONVEYANCE OF PATHOGENIC
BREATHING AGENTS WITH MULE DUCKS AND ITS CLINICAL CONSEQUENCES
ABSTRACT :
The breathing syndromes are a pathological chief characteristic of the breeding of mule
ducks. Several bacterial (Pasteurella multocida, Riemerella anatipestifer, Escherichia coli,
Ornithobacterium rhinotracheale, Mycoplasma sp., Chlamydophila psittaci), fungus
(Aspergillus fumigatus), or infectious agents (duck Metapneumovirus) may be involved in
those syndromes. However, those germs may as well form the subject of a sound conveyance
and we can’t make a definitive statement of the respective part of those agents and of the
factors of technical risks yet. The object of this work is to assess the conveyance of those
agents on mule ducks on poultry farms and to set up connections between the conveyance and
the clinical status of those animals. A witness case study was made between February and
June 2004 in the South West of France. Eighteen sets of mule ducks were taken blood
samples from and tested. If the conveyance of breathing pathogenic agents seem important
and polymorphous, no causal relation statistically significant between this conveyance and the
clinical status of those animals can have been set up.
KEY WORDS : RESPIRATORY DISEASE / MULE DUCK / CHLAMYDOPHILA /
RIEMERELLA / PASTEURELLA / ORNITHOBACTERIUM / ESCHERICHIA COLI /
MYCOPLASMA / ASPERGILLUS / METAPNEUMOVIRUS

MINISTERE DE L'AGRICULTURE ET DE LA PECHE
ECOLE NATIONALE VETERINAIRE DE TOULOUSE
Directeur : M. P. DESNOYERS
Directeurs honoraires……. : M. R. FLORIO
M. J. FERNEY
M. G. VAN HAVERBEKE
Professeurs honoraires….. : M. A. BRIZARD
M. L. FALIU
M. C. LABIE
M. C. PAVAUX
M. F. LESCURE
M. A. RICO
M. A. CAZIEUX
Mme V. BURGAT
M. D. GRIESS
M. J. CHANTAL
M. J.-F. GUELFI
M. M. EECKHOUTTE
PROFESSEURS CLASSE EXCEPTIONNELLE
M. CABANIE Paul, Histologie, Anatomie pathologique
M. DARRE Roland, Productions animales
M. DORCHIES Philippe, Parasitologie et Maladies Parasitaires
M. TOUTAIN Pierre-Louis, Physiologie et Thérapeutique
PROFESSEURS 1ère CLASSE
M. AUTEFAGE André, Pathologie chirurgicale
M. BODIN ROZAT DE MANDRES NEGRE Guy, Pathologie générale, Microbiologie, Immunologie
M. BRAUN Jean-Pierre, Physique et Chimie biologiques et médicales
M. DELVERDIER Maxence, Histologie, Anatomie pathologique
M. EUZEBY Jean, Pathologie générale, Microbiologie, Immunologie
M. FRANC Michel, Parasitologie et Maladies Parasitaires
M. MARTINEAU Guy-Pierre, Pathologie médicale du Bétail et des Animaux de basse-cour
M. MILON Alain, Pathologie générale, Microbiologie, Immunologie
M. PETIT Claude, Pharmacie et Toxicologie
M. REGNIER Alain, Physiopathologie oculaire
M. SAUTET Jean, Anatomie
M. SCHELCHER François, Pathologie médicale du Bétail et des Animaux de basse-cour
PROFESSEURS 2e CLASSE
Mme BENARD Geneviève, Hygiène et Industrie des Denrées Alimentaires d'Origine Animale
M. BERTHELOT Xavier, Pathologie de la Reproduction
M. CONCORDET Didier, Mathématiques, Statistiques, Modélisation
M. CORPET Denis, Science de l'Aliment et Technologies dans les industries agro-alimentaires
M. DUCOS Alain, Zootechnie
M. DUCOS DE LAHITTE Jacques, Parasitologie et Maladies parasitaires
M. ENJALBERT Francis, Alimentation
M. GUERRE Philippe, Pharmacie et Toxicologie
Mme KOLF-CLAUW Martine, Pharmacie -Toxicologie
M. LEFEBVRE Hervé, Physiologie et Thérapeutique
M. LIGNEREUX Yves, Anatomie
M. PICAVET Dominique, Pathologie infectieuse
PROFESSEUR ASSOCIE
M. HENROTEAUX Marc, Médecine des carnivores
INGENIEUR DE RECHERCHES
M. TAMZALI Youssef, Responsable Clinique équine
PROFESSEURS CERTIFIES DE L’ENSEIGNEMENT AGRICOLE
Mme MICHAUD Françoise, Professeur d'Anglais
M. SEVERAC Benoît, Professeur d'Anglais

MAÎTRE DE CONFERENCES HORS CLASSE
M. JOUGLAR Jean-Yves, Pathologie médicale du Bétail et des Animaux de basse-cour
MAÎTRE DE CONFERENCES CLASSE NORMALE
M. ASIMUS Erik, Pathologie chirurgicale
M. BAILLY Jean-Denis, Hygiène et Industrie des Denrées Alimentaires d'Origine Animale
M. BERGONIER Dominique, Pathologie de la Reproduction
M. BERTAGNOLI Stéphane, Pathologie infectieuse
Mme BOUCRAUT-BARALON Corine, Pathologie infectieuse
Mlle BOULLIER Séverine, Immunologie générale et médicale
Mme BOURGES-ABELLA Nathalie, Histologie, Anatomie pathologique
M. BOUSQUET-MELOU Alain, Physiologie et Thérapeutique
Mme BRET-BENNIS Lydie, Physique et Chimie biologiques et médicales
M. BRUGERE Hubert, Hygiène et Industrie des Denrées Alimentaires d'Origine Animale
Mlle CADIERGUES Marie-Christine, Dermatologie
Mme CAMUS-BOUCLAINVILLE Christelle, Biologie cellulaire et moléculaire
Mme COLLARD-MEYNAUD Patricia, Pathologie chirurgicale
Mlle DIQUELOU Armelle, Pathologie médicale des Equidés et des Carnivores
M. DOSSIN Olivier, Pathologie médicale des Equidés et des Carnivores
M. FOUCRAS Gilles, Pathologie du bétail
Mme GAYRARD-TROY Véronique, Physiologie de la Reproduction, Endocrinologie
M. GUERIN Jean-Luc, Productions animales
Mme HAGEN-PICARD Nicole, Pathologie de la Reproduction
M. JACQUIET Philippe, Parasitologie et Maladies Parasitaires
M. JAEG Jean-Philippe, Pharmacie et Toxicologie
M. LYAZRHI Faouzi, Statistiques biologiques et Mathématiques
M. MARENDA Marc, Pathologie de la reproduction
M. MATHON Didier, Pathologie chirurgicale
Mme MESSUD-PETIT Frédérique, Pathologie infectieuse
M. MEYER Gilles, Pathologie des ruminants
M. MONNEREAU Laurent, Anatomie, Embryologie
Mme PRIYMENKO Nathalie, Alimentation
Mme RAYMOND-LETRON Isabelle, Anatomie pathologique
M. SANS Pierre, Productions animales
Mlle TRUMEL Catherine, Pathologie médicale des Equidés et Carnivores
M. VERWAERDE Patrick, Anesthésie, Réanimation
MAÎTRE DE CONFERENCES CONTRACTUELS
M. CASSARD Hervé, Pathologie du bétail
N. DESMAIZIERES Louis-Marie, Clinique équine
M. LEON Olivier, Elevage et santé en productions avicoles et porcines
MAÎTRE DE CONFERENCES ASSOCIE
M. REYNOLDS Brice, Pathologie médicale des Equidés et Carnivores
ASSISTANTS D'ENSEIGNEMENT ET DE RECHERCHE CONTRACTUELS
M. CORBIERE Fabien, Pathologie des ruminants
Mlle LACROUX Caroline, Anatomie pathologique des animaux de rente
Mme MEYNADIER-TROEGELER Annabelle, Alimentation
M. MOGICATO Giovanni, Anatomie, Imagerie médicale
Mlle PALIERNE Sophie, Chirurgie des animaux de compagnie

5
A notre président de thèse,
Monsieur le Professeur Henri DABERNAT
Professeur des Universités
Praticien Hospitalier
Bactériologie-Virologie.
Qui nous a fait l’honneur d’accepter la présidence de notre jury de thèse.
Qu’il nous soit permis de vous adresser notre hommage respectueux.
A notre jury de thèse,
Monsieur le Docteur Jean-Luc GUERIN
Maître de conférence de l’Ecole Nationale Vétérinaire de Toulouse
Productions animales. Elevage et Santé Avicole et Cunicole.
Qui a inspiré et guidé notre travail et nous a soutenu pendant la réalisation de cette thèse.
Qu’il trouve l’expression de notre sincère gratitude.
Monsieur le Professeur Guy BODIN ROZAT DE MANDRES NEGRE
Professeur de l’Ecole Nationale Vétérinaire de Toulouse
Pathologie générale. Microbiologie. Immunologie.
Qui nous a fait l’honneur de participer à notre jury de thèse.
Qu’il veuille bien accepter le témoignage de notre profond respect.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
1
/
86
100%