2.3 Anatomie - Manuel de l`évolution biologique

2.3 - L'anatomie comparée
Le principe de l'ascendance commune implique que tous les organismes ont
pour origine des ancêtres communs (voir la section 4.2 : « Le darwinisme ») ;
comparer les structures entre les différents groupes fossiles permet de saisir les voies
suivies par l'évolution. L’anatomie comparée s’est dotée de plusieurs principes pour
dégager les caractéristiques d’un animal afin de le situer par rapport à un milieu, à
d’autres espèces ou à un niveau taxinomique.
Le principe de la « corrélation des organes », encore appelé « loi de corrélation » ou
bien « loi de coexistence des organes », est l’un des plus importants. Établi en 1795
dans un mémoire d’Étienne GEOFFROY SAINT-HILAIRE et de Georges CUVIER, il affirme
qu’un organe ne peut changer sans en affecter d’autres. Par conséquent, chaque
organe, étudié séparément, peut donner des informations sur d’autres. G. CUVIER
utilisera avec succès ce principe dans la reconstruction du Paleotherium des gypses
de Montmartre.
Le principe des connexions de É. GEOFFROY SAINT-HILAIRE, qui stipule que les connexions
entre les organes sont des invariants, autorise la reconnaissance des homologies (voir
la section 2.2.3 : « La méthode cladistique, la reconnaissance des homologies »).
Un troisième principe, fréquemment utilisé, lie les structures organiques des animaux à
leur milieu et à leur mode de vie. On parlerait aujourd’hui de la « structure-fonction »
des organes. LAMARCK et É. GEOFFROY SAINT-HILAIRE en ont été les premiers utilisateurs.
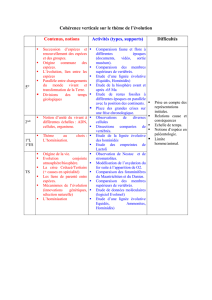
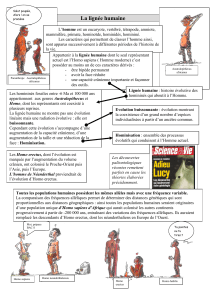
Les Hominoïdes offrent l'exemple d'une lignée reconstituée à partir de l'analyse
et de la comparaison de pièces squelettiques fossiles. Leur étude, qui repose sur des
mesures précises, a permis de retracer leur morphologie générale et de reconstituer
parfois certains de leurs comportements, par exemple la taille des outils. Une datation
précise des fossiles s'avère indispensable, car la lignée humaine a évolué rapidement.
Malheureusement, les sédiments quaternaires, souvent remaniés dans quelques
régions, sont difficiles à dater et les filiations sont régulièrement révisées ; les
datations de fossiles africains, demeurés enfouis, sont parfois très fiables, car les
sédiments du Rift africain, épais de 1 200 m, sont peu remaniés et les accidents
volcaniques offrent de bons repères chronologiques. Les préhistoriens s'accordent
néanmoins sur les grands traits de la lignée proposée.
120

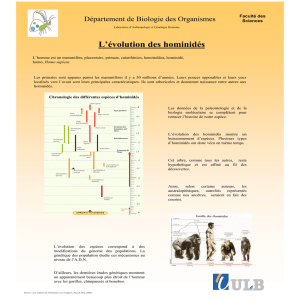
2.3.1 - L'anatomie comparée et la filiation des Hominidés
Pour suivre les discussions exposées dans cette section, un aperçu de la
classification des Primates (fig. 2.24) est nécessaire. Cette classification utilise des
critères morphologiques classiques, mais aussi des données biochimiques et
chromosomiques.
Une classification admise comme traditionnelle (fig. 2.24-A) montre que les
Hominoïdes réunissent les Singes anthropoïdes (Grands Singes), les Hommes et les
ancêtres communs aux deux groupes. Pour les cladistes, elle est devenue obsolète,
car la famille des Pongidés est paraphylétique : les Gibbons, les Orangs-outans, les
121

Gorilles et les Chimpanzés n’ont pas un ancêtre commun direct, le taxon est artificiel.
En revanche, les évolutionnistes admettent cette paraphylie, car ils considèrent non
pas des clades (ensembles de taxons ayant une filiation directe) mais des grades
(taxons ayant atteint un même niveau évolutif) : la coupure Pongidés-Homme est
justifiée.
La classification de M. GOODMAN (1962) est la première à proposer une proche parenté,
confirmée ensuite par la biologie moléculaire, des Orangs-outans, des Gorilles, des
Chimpanzés et de l’Homme (fig. 2.24-B). C’est pourquoi ces quatre genres sont réunis
dans la sous-famille des Homininae .
Mais l’analyse cladistique réfute la famille des Hominidés définie par M. GOODMAN, qui
est paraphylétique ; cette famille, qui comprend les genres Sahelanthropus, Orrorin,
Ardipithecus, Paranthropus, Australopithecus et Homo, concerne uniquement la lignée
hominienne, c’est-à-dire celle qui conduit directement à l’Homme. La classification
cladistique des Hominidés (fig. 2.24-C), confirmée elle aussi par la biologie
moléculaire, est acceptée par la grande majorité des chercheurs. Mis à part les noms
des taxons, les classifications des figures 2.24-B et 2.24-C sont identiques.
Les caractères morphologiques utilisés
Pour reconstituer la filiation des Hominidés, les chercheurs s'appuient sur des
caractères morphologiques, squelettiques et dentaires. L'évolution de cette lignée se
distingue par trois traits principaux : l'acquisition de la station verticale et de la
locomotion bipède, la réduction de la denture et, enfin, le fort développement de
l'encéphale.
- La verticalité
Son acquisition entraîne une restructuration fondamentale du bassin, ainsi que
celle des pieds. La bipédie qui en résulte est avant tout liée aux remaniements du
bassin. La figure 2.25 montre quelles sont les mesures effectuées chez les
Hominoïdes.
122

Au cours de la phylogenèse, la stabilité de la bipédie s'améliore, quand la largeur et le
rapport hauteur/largeur du bassin diminuent. Enfin, on a reconstitué à la fois l'ordre
d'apparition des grands caractères spécifiques à cette lignée et les parentés entre les
différents genres.
L’acquisition de la bipédie se manifeste également par un déplacement du trou
occipital vers l’avant, déplacement d’autant plus prononcé que la bipédie est parfaite.
123

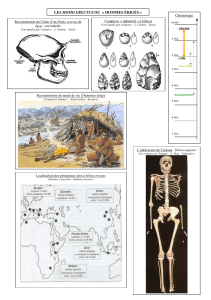
Sa date d’apparition recule de plus en plus. En 1976, la découverte de trace de pas
fossilisée à Laetoli (Tanzanie) l’a située à 3,7 Ma probablement chez les
Australopithèques ; puis, en 2000, celle d’Orrorin tugenensis la place à 6 Ma ; et enfin,
en 2001, celle de Sahelanthropus tchadensis (Toumaï) la remonte à 7 Ma.
- La denture
La réduction du système dentaire est l'une des caractéristiques principales de
l'hominisation. Sont pris en compte d'autres critères tels que la hauteur, la largeur des
dents, le nombre de tubercules (indice d'un régime omnivore) de la surface broyeuse
des prémolaires et des molaires, la taille décroissante des molaires d’arrière en avant
chez les Hominidés (celles situées au fond de la mâchoire sont généralement les plus
grandes), alors que chez les Pongoïdes leur taille est croissante. La forme de l'arc
dentaire fournit également une indication sur le degré d'hominisation : les Singes ont
un arc dentaire fermé en forme de U ; en revanche dans la lignée humaine, il tend à
s'évaser de plus en plus. La mandibule puissante et robuste chez les Hominidés
primitifs s'allège progressivement, tandis que le menton, absent chez les Pongoïdes et
les Hommes primitifs, se développe.
- La cérébralisation
Elle s'accompagne d'une modification des supports osseux : crâne,
superstructures crâniennes (torus ou bourrelet sus-orbitaire, crête sagittale, apophyse
mastoïde et torus ou chignon occipital), et mâchoires. Un tableau comparatif (fig. 2.26)
présente les mesures crâniennes de plusieurs espèces d'Hominidés.
124
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
1
/
21
100%