partie 7 - CHUPS – Jussieu

Pi
+
ADP
Pi
+
ADP
Resynthèse
de l'ATP
Pmax
V0
Commande nerveuse
Activation musculaire
(potentiels d'action)
Relargage du Calcium
(reticulum sarcoplasmique)
ATPATP
F0
Hydrolyse de
l'ATP
( Myosine-ATPase)
Phénomènes
physiologiques
Phénomènes
biochimiques
Phénomènes
mécaniques
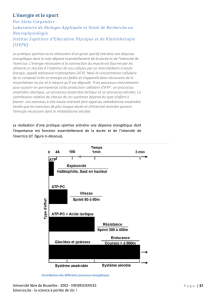
Les -1
(environ 4 à 7 mmoles.l ) et
ne permettent en théorie que
quelques contractions.
réserves d'ATP sont très
faibles
Le muscle dispose de
non directement
utilisable, sous forme de
phosphocréatine, de glucides, de
lipides.
réserves
d'énergie
La poursuite de
l'exercice exige
la
.
resynthèse de
l'ATP
?

Pi
+
ADP
Pi
+
ADP
Resynthèse
de l'ATP
Pmax
V0
Métabolisme
aérobie
Métabolisme
anaérobie
lactique
Métabolisme
anaérobie
alactique
Commande nerveuse
Activation musculaire
(potentiels d'action)
Relargage du Calcium
(reticulum sarcoplasmique)
ATPATP
F0
Hydrolyse de
l'ATP
( Myosine-ATPase)
Phénomènes
physiologiques
Phénomènes
biochimiques
Phénomènes
mécaniques
Trois métabolismes
énergétiques permettent la
resynthèse de l’ATP au cours
de l’exercice.
?

Bioénergétique et Entraînement
Dans le modèle bioénergétique de la performance, il est
classique de caractériser un métabolisme énergétique par
son inertie, sa puissance maximale et sa capacité de
travail.
D’après ce modèle bioénergétique, les entraînements
peuvent être différenciés en fonction de leur action
préférentielle sur le développement de la puissance et de la
capacité maximales des trois métabolismes énergétiques.

Caractéristique du métabolisme énergétique
Inertie
Puissance maximale
Capacité de travail
: lenteur d’entrée en action, c’est-à-dire temps
nécessaire pour atteindre un pourcentage élevée de la
puissance requise.
: débit maximal d’énergie, c’est-à-dire
débit d’ATP.
: quantité totale d’énergie produite, c’est-
à-dire quantité totale d’ATP restaurée.

DragsterDragster
Métabolisme anaérobie alactique
Inertie : extrêmement faible ou nulle
Inertie : extrêmement faible ou nulle
Puissance : très élevée
Capacité de travail : très faible
Démarrage très court
Puissance très élevée
Distance parcourue très faible
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
1
/
149
100%